
Жизнь человека по меркам Вселенной — всего лишь мгновение, а по меркам мухи-однодневки — целая вечность. Для нас же оценка продолжительности нашего собственного жизненного пути осложнена событиями, которые происходят в процессе, людьми, которых мы встречаем и с которыми расстаемся, эмоциями, которые испытываем. Ведь чем сложнее мозг существа, тем сложнее его самосознание. Однако любой путь определяется не только его насыщенностью, но и фактом того, что он рано или поздно заканчивается. Люди многие века пытаются ответить на вопрос, что лежит за гранью жизни, и четкого безапелляционного ответа нет ни у кого. Тем не менее «до» остается не менее таинственным и загадочным, чем «после». Ученые из Американского музея естественной истории (Нью-Йорк, США) провели исследование, в котором описывается возможный вариант зарождения органических молекул миллиарды лет тому назад. Что могло послужить началом жизни на планете, где это произошло, и как эти знания могут помочь в понимание нашего мира здесь и сейчас? Ответы на эти вопросы сокрыты в докладе ученых. Поехали.
Основа исследования
Как уже было сказано ранее, достоверно неизвестно, что ждет человека или любой другой живой организм после того, как его тело теряет жизнеспособность. Этим вопросом задаются физики, биологи, теологи и философы. У все у них есть ответы, каждый из которых имеет право на существование, однако это лишь теории, которые эмпирически подтвердить по понятным причинам пока никому не удавалось.
Что касается того, что было до начала всего, то этот вопрос такой же сложный и неоднозначный. Теория большого взрыва подарила нам идею зарождения Вселенной, но остаются вопросы касательного того, что было до него. Эволюционная теория Дарвина помогла понять как между собой взаимосвязаны виды на нашей планете, как они эволюционировали, как одно преобразовалось в другое. Но и тут возникают вопросы: что или кто был первым, почему жизнь зародилась, при каких обстоятельствах, случайно ли было это событие или это чей-то великий замысел. Вопросы на века, не иначе.
Тем не менее, знания, которыми обладает современный человек, могут послужить инструментом в построении цепочки событий, которые привели к зарождению жизни. Нам известно, что основополагающими элементами в зарождении и поддержании жизни являются водород, азот и кислород. В современной жизни большинство органических молекул образуются в результате восстановления углекислого газа (CO2) посредством нескольких путей «фиксации углерода» (например, фотосинтез в растениях). Но большинство этих путей либо требует энергии от клетки, либо они появились относительно поздно. Возникает вопрос — что было до этого?
По мнению ученых одним из путей образования органики могло быть восстановление CO2 с помощью H2. Геологические исследования показывают, что СО2 находился в относительно высоких концентрациях в океане во время катархея*, тогда как H2 был продуктом множественных процессов в земной коре и выделялся наружу за счет гидротермальных источников.
Катархей* — геологический эон (период времени), продлившийся первые 600 миллионов лет существования Земли.Следовательно, на стыке двух сред (океана и Земной коры) между двумя растворенными газами возникала реакция, которая приводила к образованию углеводородов, сыгравших в последствии важную роль в переходе от геохимии к биохимии.
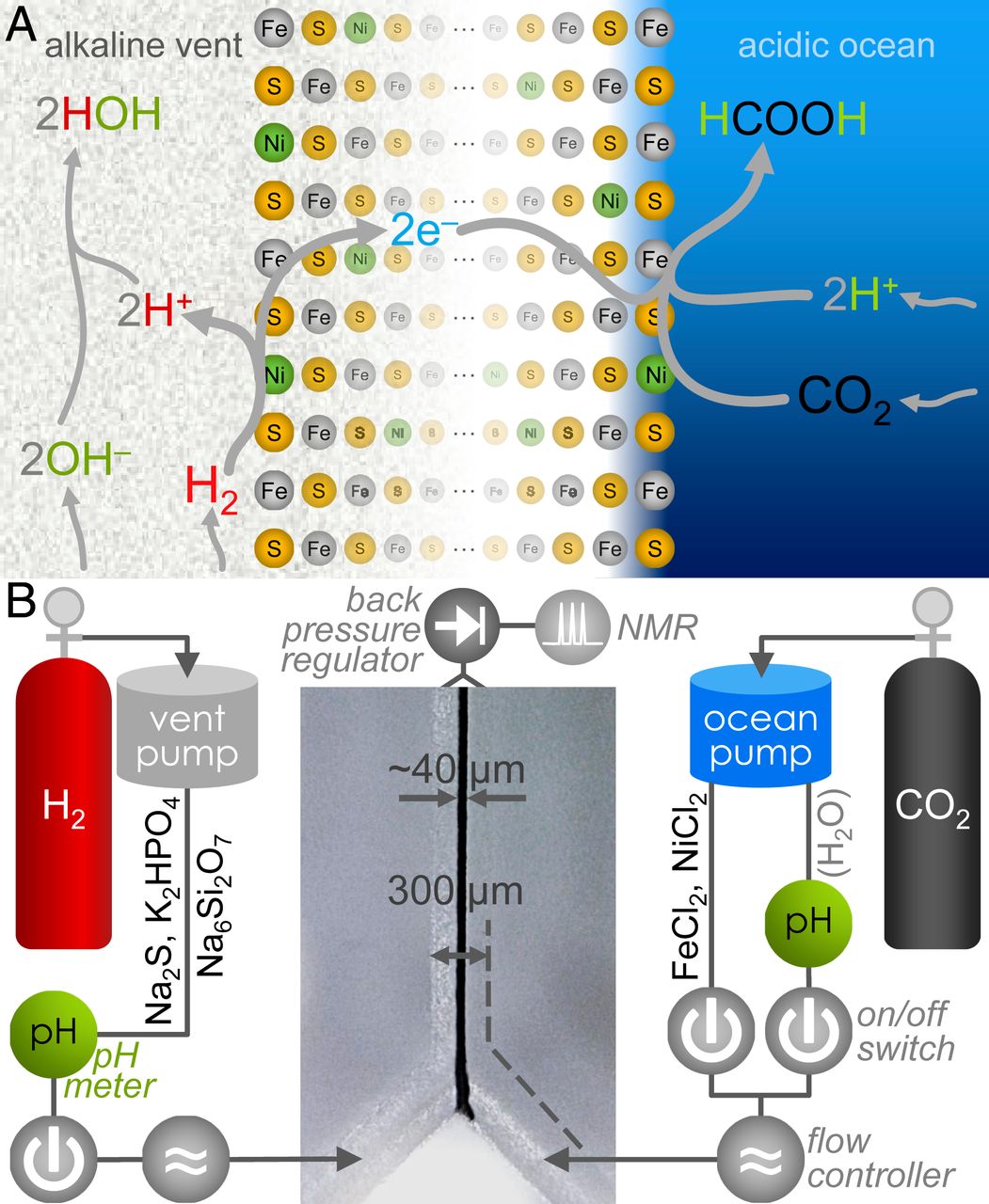
Изображение №1
В стандартных условиях (1 ат, 25 °C, pH 7) реакция между CO2 и H2 с образованием формиата (HCOO–) термодинамически неблагоприятна с ΔG0’ = + 3.5 кДж/моль. Однако в древних щелочных источниках (1А) H2 присутствовал в богатых ОН– водах гидротермального источника, что способствовало его окислению в воду. При этом CO2 был бы растворен в относительно кислом океане, что облегчило протонирование в его восстановлении до HCOO–.
С помощью минералов Fe(Ni)S, осажденных на интерфейсе между океаном и корой, градиента pH более трех единиц должно было быть достаточно для увеличения жизнеспособности реакции на ~ 180 мВ, что делает ее благоприятной для образования органики.
После образования формиат обладал бы достаточным абиотическим химическим потенциалом. К примеру, известно, что формильные группы образуют промежуточные соединения восстановительного цикла трикарбоновых кислот* и восстановительного Ацетил-КоА пути*, предполагая возможный путь развития биологического метаболизма.
Восстановительный цикл трикарбоновых кислот* — череда химических реакций синтеза органики из диоксида углерода и воды.
Восстановительный Ацетил-КоА путь (путь Вуда-Льюнгдаля или WL-путь)* — цепочка биохимических реакций, необходимых для фиксации СО2 и для получения энергии.Другая теория предполагает, что при нагревании в присутствии аммиака, который также является предполагаемым компонентом щелочных вод, из формиата образуется формамид [HC(O)NH2] — высокореакционная молекула, являющаяся краеугольным камнем одной из теорий возникновения жизни (Formamide and the origin of life). Дальнейшая реакция этой смеси дает цианистый водород (HCN), который также является основой еще одной теории образования органики (Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism). В свою очередь, дегидратация формиата приводит к образованию монооксида углерода (CO) (Activated Acetic Acid by Carbon Fixation on (Fe,Ni)S Under Primordial Conditions). Как видно, теорий происхождения органических соединений немало, и каждая из них имеет вполне логичные объяснения.
Несмотря на то, что на ранней Земле существовало несколько источников восстановленного углерода и множество вероятных сред, в которых могли бы находиться богатые химические вещества, описанный выше щелочной гидротермальный сценарий куда более интересен для ученых ввиду его сходства с WL-путем фиксации углерода.
Дополнительным аргументом в пользу потенциальной значимости щелочного гидротермального сценария образования органики является факт того, что WL-процесс это единственный из шести известных биологических путей фиксации углерода, который высвобождает энергию в целом, а не потребляет ее, а его вариации присутствуют у существующих представителей обеих архей (метаногены*) и бактерии (ацетогены*).
Метаногены* — археи, образующие метан как побочный продукт метаболизма в бескислородных условиях.
Ацетогены* — бактерии, которые выделяют ацетат (CH3COO–) в качестве конечного продукта анаэробного дыхания или гомоацетатного брожения.Первым шагом на этом пути является восстановление CO2 с помощью H2 с образованием формиата (HCOO– или его дегидратированный электронный эквивалент, т.е. CO).
Эта реакция носит эндергонический* характер, поэтому некоторые представители как архей, так и бактерий используют либо бифуркацию электронов*, либо хемиосмос* через клеточную мембрану, чтобы привести в действие это процесс.
Эндергонические реакции* — химические реакции, требующие энергии извне для их протекания.
Бифуркация электронов* — механизм разделения электронов в окислительно-восстановительной реакции.
Хемиосмос* — преобразование энергии цепи переноса электронов в энергию АТФ (аденозинтрифосфат).Однако в отсутствие механизмов клеточного объединения, таких как бифуркация электронов или хемиосмос, эта первая эндергоническая стадия является ключевым энергетическим узким местом в WL-пути и остается главным открытым вопросом в исследованиях происхождения биологической фиксации углерода.
В данном исследовании ученые демонстрируют абиотическое косвенное восстановление CO2 до HCOO– с помощью H2, вызванное микрофлюидным градиентом pH в осадках Fe(Ni)S, посредством механизма, который напоминает поток разделенных электронов пути WL.
Результаты исследования
В первую очередь был подготовлен лабораторный эквивалент щелочной гидротермальной среды с имитацией интерфейса между корой Земли и водами океана. Щелочная составляющая включала в себя Na2S (100 мМ), K2HPO4 (10 мМ) и Na2Si3O7 (10 мМ) в деаэрированной воде. Аналог океана включал в себя FeCl2 (50 мМ) и NiCl2 (5 мМ). Обе жидкости были подведены к Y-образному боросиликатному микрофлюидному реактору (1B).
Окружающее давление H2 и CO2 оказалось недостаточным для сокращения выбросов CO2, поэтому вместо попытки растворения любого газа путем барботирования* перед реакцией было решено использовать микрожидкостные насосы, работающие от давления газа.
Барботирование (барботаж)* — процесс пропускания газа через слой жидкости.Щелочная жидкость вытеснялась H2 при давлении 1.5 бар, а аналог океана выталкивали CO2 при том же давлении.
Каждый цикл реактора был разделен на две последовательные стадии: первая — для осаждения осадков Fe(Ni)S на стыке (на интерфейсе) двух жидкостей; вторая («постосаждение») — для попытки создания реакции между CO2 и H2 (или других реагентов).
В результате взаимодействия щелочной жидкости и океанического аналога в течение 15-60 секунд на стадии выпадения осадков образовался осадок шириной от 30 до 60 мкм на интерфейсе между двух жидкостей, видимый под цифровым оптическим микроскопом (в центре на 1В). Удаление металлов со стороны аналога океана после выпадения осадков предотвратило увеличение осадка до критического значения перекрытия канала реактора.
После образования осадка и для предотвращения засорения микрофлюидных каналов дальнейшим осаждением на второй стадии океаническая жидкость была переключена на чистую деаэрированную воду, вытесненную CO2 (справа на 1B). При этом аналог щелочной жидкости оставался прежним с Na2S, K2HPO4 и Na2Si3O7, выталкиваемыми H2.
Далее был определен уровень pH поступающих жидкостей в точке входа: аналог океана — pH 3.9, щелочные воды — pH 12.3. При скорости потока 5 мкл/мин для каждого входа время пребывания жидкостей в центральном канале составляло ~ 3.3 с, поэтому системе было позволено работать не менее 2 минут перед сбором выходных данных. Далее был собран общий выход реактора (смесь жидкостей), который был проанализирован с помощью ЯМР-спектроскопии. Анализ показал, что среднее значение концентрации HCOO– составило 1.5 мкМ.

Таблица №1: результаты экспериментов.
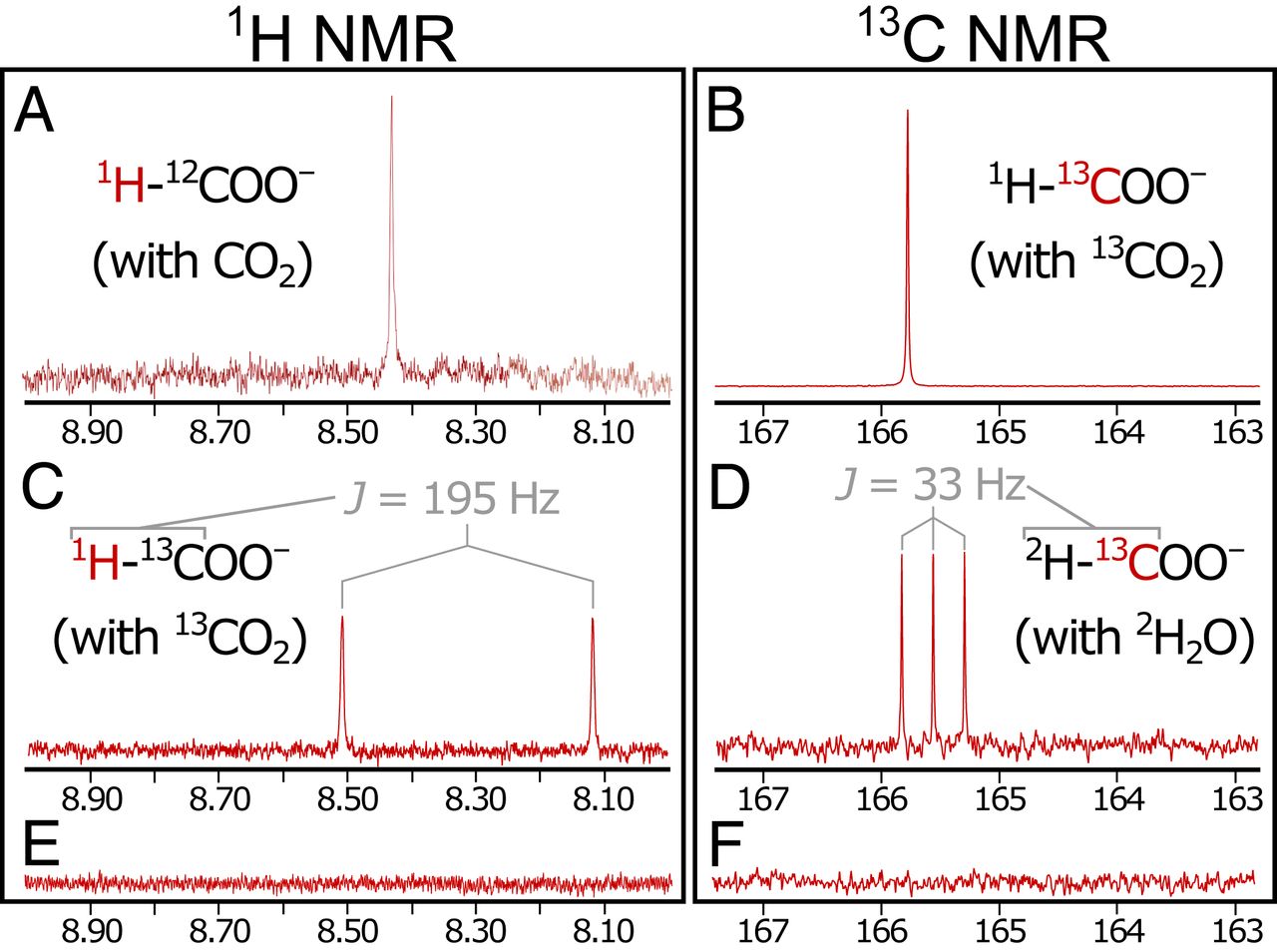
Изображение №2
Синглетные пики в спектрах 1H ЯМР (8.42 ppm (частей на миллион); 2A) и 13C ЯМР (165.8 ppm) соответствуют образцам чистой (> 98%) муравьиной кислоты. Выполнение стадий осаждения и реакции с изотопно обогащенным (99% 13C) 13CO2 (эксперимент 2) дало более сильный синглет в спектре 13C (165.8 ppm; 2B) и ожидаемое расщепление синглета формила на дублет (сигнал, расщепленный на два пика) в спектре 1H (J = 195 Гц) за счет взаимодействия 1H – 13C в формильной группе (2С).
Как оказалось, H2 необходим для сокращения выбросов CO2. С жидкостью на стороне выпускного отверстия, управляемой N2 вместо H2 (т.е. в отсутствии H2 как во время, так и после осаждения), продуктов восстановления не было обнаружено (эксперимент 3; 2E и 2F).
Для более детального понимания происходящего процесса были выполнены дополнительные эксперименты по маркировке дейтерием (2H или D) (эксперименты 4 и 5), используя изотопные варианты на протяжении всех экспериментов.
Независимо от того, использовался ли немаркированный H2 (эксперимент 1) или D2 (эксперимент 4) для управления насосом на стороне щелочной жидкости, наблюдались исключительно неизотопно отмеченный HCOO– в жидкостном выходе. Это наблюдение позволяет предположить, что сокращение выбросов CO2 может происходить исключительно на стороне океана.
И наоборот, с D2O, используемым вместо обычного H2O на стороне океана, и с немаркированным H2, приводящим в действие насос на стороне щелочной жидкости (эксперимент 5), был обнаружен исключительно дейтерированный формиат (DCOO–), о чем свидетельствует триплет в 13C ЯМР (J = 33 Гц) и отсутствие каких-либо других заметных пиков (2D). Это дополнительно подтверждает, что сокращение CO2 соответствует изотопному составу на стороне океана, а не на стороне земной коры.
На следующем этапе исследования была проверена роль градиента pH моделируемой подводной щелочной гидротермальной системы. Успешные сокращения CO2, представленные в таблице 1, происходили при pH аналога океана 3.9 и pH аналога сброса 12.3.
При смешивании этот начальный ΔpH, равный 8.4 единиц, неизбежно снизился бы, но градиенты pH, составляющие несколько единиц, успешно сохраняются с течением времени в микрожидкостных масштабах, особенно в присутствии осадка на интерфейсе.
Необходимо было понять, требуется ли такой градиент pH в системе восстановления для облегчения окисления H2 на щелочной стороне и восстановления CO2 на кислотной стороне (1A). После осаждения в тех же условиях, что и для эксперимента 1, оценивались эффекты различных уровней pH и состава каждой из двух жидкостей (таблица №2). Замена имитатора щелочного источника чистым H2O, управляемым H2, не дала результата (таблица 2, эксперимент 6).
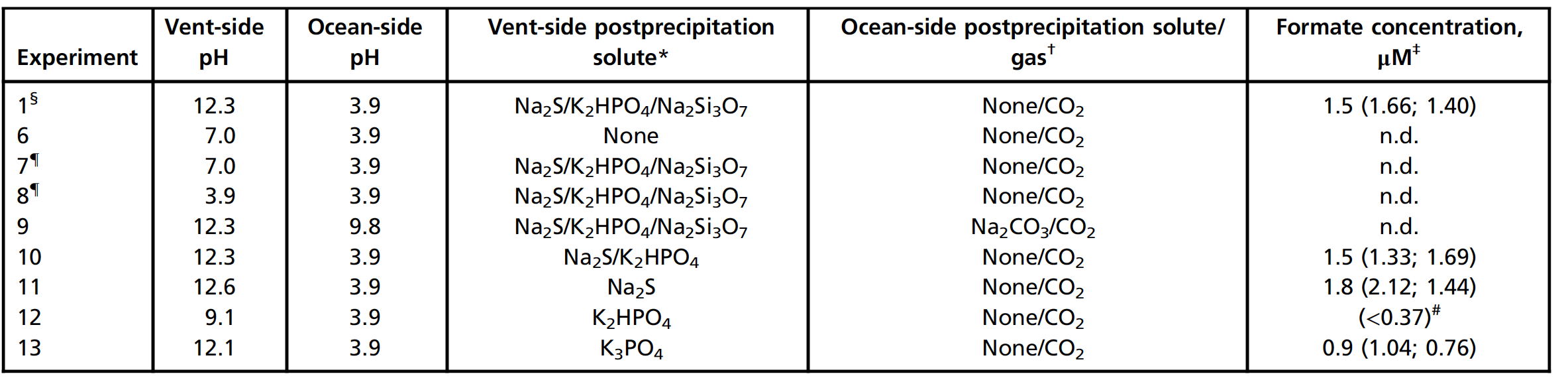
Таблица №2: результаты экспериментов с различным значением рН.
Аналогичным образом, подкисление жидкости аналога щелочного источника с помощью HCl до pH 3.9 и pH 7.0 не привело к образованию формиата (эксперименты 7 и 8).
Добавление 100 мМ Na2CO3 в океаническую жидкость при одновременном использовании CO2 в качестве движущего газа (эксперимент 9) повысило pH океана до 9.8, и в этих условиях продукт не был обнаружен. Удаление силиката со стороны источника после осаждения все еще давало формиат (эксперимент 10), как и удаление силиката и фосфата при наличии только Na2S (эксперимент 11).
Имея только K2HPO4 после осаждения на стороне щелочного источника, были обнаружены лишь остаточные количества формиата (ниже предела количественного определения в 0.37 мкМ), возможно, из-за недостаточно щелочного pH 9.1 (эксперимент 12). А вот более щелочной K3PO4 повысил pH до 12.1 и привел к образованию значительно большего количества формиата (эксперимент 13).
Ученые заявляют, что не могут полностью исключить вероятность того, что связанный с осадком сульфид действует как восстановитель в дополнение к H2. Однако вышеописанные результаты одновременно подтверждают роль градиента pH и показывают, что непрерывная подача водного сульфида в системе не требуется.
Удаление Ni из осадочной жидкости океана (эксперимент 14) привело к образованию лишь небольшого количества формиата. И наоборот, замена Fe, чтобы оставить Ni в качестве единственного металла в осадочной жидкости океана (NiCl2, 55 мМ; эксперимент 15), дало 1.4 мкМ формиата, что указывает на решающую роль Ni в составе осадков.
Удаление FeCl2 и NiCl2 из океанической жидкости, как и ожидалось, не привело к образованию определяемого формиата и осадка (эксперимент 16).
Самым подходящим объяснением происходящего ученые считают именно электрохимический процесс (1А), однако существует несколько альтернативных механизмов сокращения выбросов CO2, связанных с окислением H2, которые при этом менее вероятны.
Один из таких механизмов можно назвать самым «простым», но и наименее биохимически гомологичным — восстановление углерода за счет прямого гидрирования (3А-3С). В таком варианте водород из H2 будет передаваться непосредственно в CO2 либо в виде атомарного водорода (классическое гидрирование) или в виде гидрида (ионное гидрирование).
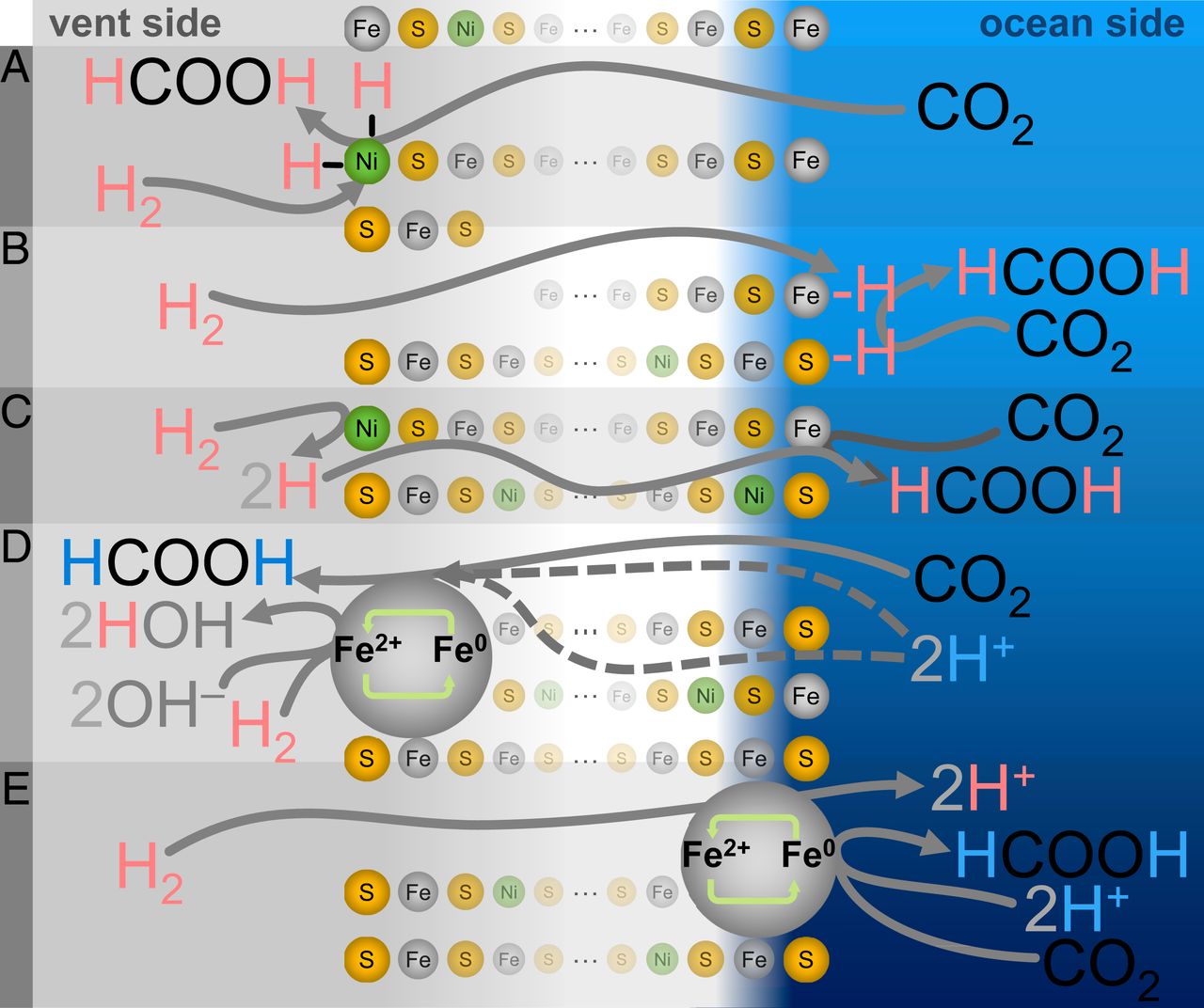
Изображение №3
Другими словами, выходной продукт в таком механизме должен соответствовать изотопной сигнатуре выделяемого газа H2/D2. Вместо этого произведенный в таком случае формиат соответствует только изотопному составу воды на стороне океана, независимо от состава газа или воды на стороне гидротермального источника.
В механизмах прямого гидрирования адсорбированные частицы водорода могут обмениваться с окружающей жидкостью, так что исходная изотопная сигнатура теряется. Однако любой такой процесс подразумевает миграцию значительного количества жидкости через осадок. Существенное перемешивание жидкостей должно было вызвать смешанный сигнал H/D формила, которого в ходе практических опытов не наблюдалось, полностью исключая гидрирование.
Другой возможный вариант заключается в том, что атомы водорода в образующемся формиате могут не происходить непосредственно из H2. Вместо этого механизм может осуществляться посредством окислительно-восстановительного цикла краевого или углового атома Fe или Ni (M2+ ⇄ M0), при котором металл сначала восстанавливается H2 (оставляя два протона для разбавления), а затем металл переносит приобретенные электроны на СО2 с сопутствующим отрывом протонов из локальной водной среды (3C-3E).
Однако такой вариант развития событий сложно сопоставим с реальными показателями рН, которые были во время опытов. Уровень рН в 3.9 достигался исключительно за счет растворения CO2 в воде. Таким образом, любые протоны со стороны океана должны происходить в результате диссоциации угольной кислоты посредством:
H2O + CO2 ⇌ H2CO3 ⇌ H+ + HCO–3
Когда же реакция проводилась с использованием D2O (эксперимент 5) в качестве растворителя на стороне океана, в выходном потоке обнаруживался исключительно DCOO–. Из этого следует, что сокращение CO2 не происходило на стороне источника, где присутствовали обычная вода (H2O) и H2.
Сценариев такого локализованного окислительно-восстановительного цикла (3D и 3E) может быть несколько, но поскольку все они требуют совместного размещения ни один из них не может предложить изотопную сигнатуру исключительно на стороне океана, которая наблюдалась во время экспериментов.
В совокупности с сильной зависимостью реакции от уровня рН эти результаты говорят о том, что восстановление СО2 происходит посредством электрохимического механизма, в котором электроны от окисления H2 на стороне щелочного источника перемещаются через осадки Fe(Ni)S в сторону CO2 на стороне океана (1A).
Вышеописанные процессы не могли бы протекать, если бы не было какого-то механизма, позволяющего активировать и поддерживать взаимодействие между океанической водой и щелочным гидротермальным источником. Кроме того возникает вопрос и касательно «живучести» сформировавшихся органических соединений, так как они могли бы попросту раствориться в океанической воде до того, как примут какую-либо биохимическую роль.
Ответом на первый вопрос может быть эффект Вентури*, вызванный повышенной пористостью структуры гидротермальных жерл. Оказавшись внутри жерла, углекислые воды океана могут вступить в реакцию с электронами, переносящимися через каталитические минералы канала гидротермального источника, а новые осадки также могут возникать и дальше по времени, когда две жидкости вступают в контакт.
Эффект Вентури* — падение давления, когда поток жидкости или газа протекает через суженную часть трубы.Моделирование данной теории показало, что в случае экспериментального реактора шириной 300 мкм действительно происходит микрожидкостное слияние двух реагентов, что и было показано в ходе практических опытов.
Ученые также отмечают, что такой эффект не ограничивается подводными щелочными жерлами и, вероятно, будет происходить в пористых гидротермальных системах в любом месте и на любой глубине, что дает возможность множеству геохимических сценариев возникновения жизни.
Стоит отметить, что микрожидкостная система восстановления СО2 с помощью H2 не является единственной. Существует также и методика, где используется одноканальная периодическая система.
За счет других минералов (Fe3Ni), более высокого давления (10 бар для H2) и более высокие температуры (100 °C), чем в проводимых опытах, система периодического действия позволяет получить куда больше формиата, а также несколько продуктов дальнейшего восстановления (ацетат, метанол и пируват). При этом скорость производства формиата (5.21 х 10-9 моль/с) на четыре порядка выше скорости, достигнутой микрожидкостной системой.
Важность периодической системы заключается в том, что ее результаты подтверждают работоспособность микрожидкостной системы. Таким образом подтверждается и теория касательно существования органических веществ в условиях бескислородных щелочных гидротермальных источников.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Все, что имеет начало, имеет и конец. Эти слова, сказанные Пифией из х/ф «Матрица», в той или иной интерпретации говорились реальными философами и учеными задолго до выхода этой кинокартины. Помимо этого, из курса школьной химии вспоминается один из основоположных принципов науки — ничто никуда не девается и ниоткуда не берется бесследно. С тем, что будет в конце, а точнее после него, разбираться человечеству придется еще очень долго. Но вот какое-то понимание того, что было в начале, уже есть.
В данном исследовании ученые описали возможный вариант формирования первых органических соединений. По их мнению этот процесс протекал на стыке океанических вод и гидротермальных источников. В ходе экспериментов удалось преобразовать СО2 в органические молекулы посредством H2 и управляемого рН.
Такой результат не только объясняет исток жизни на нашей планете, но и может быть использован в разработке инструментов для снижения выбросов CO2, что является весьма удручающей проблемой современного мира. Помимо прочего, понимание того, как органика появилась на Земле, позволяет строить более рентабельные теории о возможном наличии оной на других планетах, подобных нашей.
Если же переключиться на более философскую волну, то можно с уверенностью сказать, что данный труд демонстрирует важность понимания прошлого для успешного формирования будущего. История полнится исследованиями, которые современники называли праздным любопытством и пустой тратой времени. Подавляющее большинство из них оказались гораздо важнее, чем кто-либо мог предположить. Вывод прост: в науке искать ответы нужно на все вопросы, какими бы глупыми на первый взгляд они ни казались.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята. :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?