
Так уж сложилось, что человеку присуще создавать ассоциации, часть которых имеет вполне прямолинейную логику, а часть — более абстрактна. К примеру, слово «зима» у многих ассоциируется с холодом, снегом, новогодними мероприятиями и некомпетентностью коммунальных служб. Если же говорить о чем-то более приятном, то слово «бабочка» ассоциируется с легкостью, нежностью и, конечно же, красочностью. И правда, у многих видов чешуекрылых крылья украшены самыми невероятными цветами и замысловатыми узорами, вызывающими недоумение у хищников и восхищение у художников и поэтов. Однако некоторые бабочки и мотыльки решили отказаться от яркого одеяния в угоду полного «ню», а именно в угоду прозрачности. Предназначение прозрачных (или частично прозрачных) крыльев ясна — камуфляж, но какова природа этой удивительной способности доселе было неясно. Посему ученые из Калифорнийского университета (США) решили провести детальный анализ структуры крыла разных «прозрачнокрылых» видов, который показал разные механизмы достижения прозрачности. В чем же секрет прозрачных крыльев, и насколько сильно они отличаются от цветных с точки зрения морфологии? Ответы на эти и другие вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Мир насекомых всегда вызывал неподдельный интерес у любознательных умов: великое множество разнообразных существ, каждое из которых обладает тем или иным уникальным функциональным биологическим приспособлением, помогающим выживать в условиях постоянной внутривидовой конкуренции и межвидовой борьбы. Бабочки, относящиеся к отряду чешуекрылых (Lepidoptera), стали объектом множества исследований, в которых ученые изучали как их удивительные крылья, так и жизненный цикл, включающий полное превращение из гусеницы в бабочку.
Ранее мы с вами уже заглянули за кулисы таинственного процесса формирования крыльев в процессе метаморфоза, протекающего внутри куколки (Магия внутри кокона: как формируются крылья будущей бабочки). Правда, это исследование касалось «цветных» бабочек, которых хоть и больше, но есть и частично прозрачные (1A-1H).

Изображение №1
Ученые, недолго думая, отнесли эту особенность к камуфляжу, что вполне логично. В случае бабочек прозрачность является результатом пропускания света видимого спектра через материал, в данном случае через хитиновую мембрану, без заметного поглощения или отражения. Уровни отражения в значительной степени определяются различиями в показателях преломления между биологическими тканями и средой, и большая разница приводит к более высокому отражению.
Прозрачность в природе достаточно часто встречается у обитателей морской среды, чему способствует близкое соответствие показателей преломления их тканей и окружающей среды, т.е. воды. Но вот прозрачность на суше встречается крайне редко. Ученые считают, что это связано со сложностью достижения этой самой прозрачности ввиду большой разницы между показателями преломления тканей наземных организмов (n = ~ 1.3-1.5) и воздуха (n = 1), что приводит к значительному поверхностному отражению.
Тем не менее некоторые организмы развили морфологические особенности, которые преодолевают проблемы наземной прозрачности, особенно в форме антиотражающих наноструктур. К примеру, еще в 60-ых ученые выявили высокоупорядоченные субволновые наноструктуры на поверхности роговицы глаз насекомых (The insect corneal nipple array). Было обнаружено, что эти структуры обычно имеют высоту ~ 150–250 нм, а расстояние между ними составляет ~ 200 нм. Это снижает отражение в широком диапазоне длин волн за счет создания более плавного градиента показателей преломления между воздухом и хитином (Light on the moth-eye corneal nipple array of butterflies). Подобные массивы наноструктур также были обнаружены на крыльях цикад, которые помогают уменьшить поверхностное отражение в видимом спектре (Design for approaching Cicada-wing reflectance in low- and high-index biomimetic nanostructures).
Как мы уже поняли, некоторые чешуекрылые обладают «прозрачными крыльями», чешуя которых претерпела изменения, позволяющие свету достигать поверхности мембраны крыла. Само крыло состоит из хитина и обладает некоторой присущей ему прозрачностью, но благодаря высокому показателю преломления хитина (n = 1.56) поверхность крыла отражает свет.

Methona confusa (слева) и Chorinea faunus (справа).
К примеру, бабочка Methona confusa имеет обнаженную мембрану крыла, на поверхности которой отсутствуют наноструктуры. В результате этого крыло несколько прозрачно, но сохраняет высокую степень отражательной способности (1А-1Е). А вот Chorinea faunus имеет мелкие распределенные чешуйки и куполообразные хитиновые нановыступы на мембране, которые создают антиотражающий эффект.
Массивы наноструктур также были найдены в прозрачных областях крыльев мотылька Cacostatia ossa, а Greta oto имеет тонкие вертикально ориентированные чешуйки, позволяющие обнажать поверхность крыла, а также наностолбики, покрывающие поверхность (1F-1J).

Greta oto
Неравномерно расположенные наностолбики Greta oto имеют случайное распределение по высоте и обеспечивают всенаправленные антиотражательные свойства (1I и 1J).
Вышеописанные виды являются примером того, что у разных бабочек, коим присуща та или иная степень прозрачности крыльев, имеется разный механизм достижения этой прозрачности. Это говорит о том, что подобная физиологическая особенность развивалась у них параллельно, т.е. имеет разные (а не общий) эволюционные корни.
Дабы заполнить пробелы в знаниях касательно прозрачности среди сухопутных организмов, ученые провели исследование формирования крыльев стеклокрылой бабочки Greta oto, которая принадлежит к разнообразной трибе (∼393 вида) преимущественно прозрачных неотропических бабочек. В ходе исследования использовались конфокальная (КЛСМ) и просвечивающая электронная микроскопия (ПРЭМ) для сравнения развития крыльев, организации цитоскелета и наноструктур поверхности мембран между прозрачными и непрозрачными областями крыльев.
С помощью химических обработок, сканирующей электронной микроскопии (СЭМ) и хромато-масс-спектрометрии (ХМС) ученые обнаружили, что наноструктуры на поверхности мембраны крыла состоят из двух слоев: нижнего слоя сосковидных наноструктур на основе хитина и верхнего слоя наностолбиков на основе воска, состоящих из преимущественно длинноцепочечных алканов*.
Алканы* — ациклические углеводороды линейного или разветвлённого строения, содержащие только простые (одинарные) связи и образующие гомологический ряд с общей формулой CnH2n+2. Примером простейшего алкана является метан (CH4).Также ученые смогли выявить роль наностолбиков в резком уменьшении отражения от поверхности крыла с помощью оптической спектроскопии и аналитического моделирования.
Результаты исследования
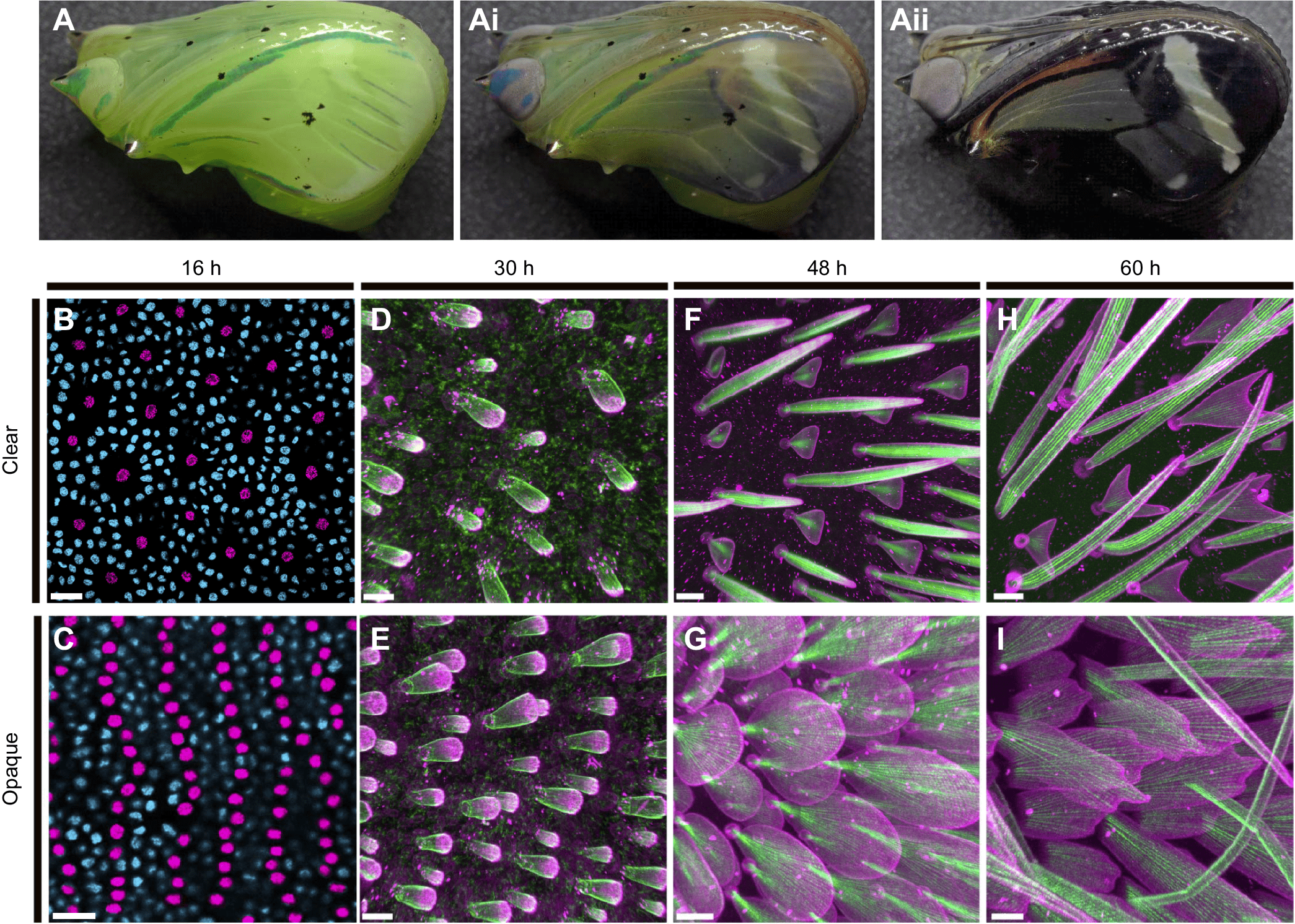
Первым делом необходимо было исследовать особенности плотности чешуек, морфологию чешуек и количество обнаженной поверхности крыла у взрослых особей Greta oto. Для упрощения этой задачи ученые сосредоточили свое внимание на двух областях на дорсальной поверхности переднего крыла: прозрачная область внутри дискоидальной ячейки (на схеме ниже отмечено синим) и непрозрачная область, состоящая в основном из черных чешуек возле поперечной жилки (выделено красным на 1F).

Схемы строения и жилкования крыла бабочек (слева сверху — переднее крыло; слева снизу — заднее крыло).
Обнаженная часть крыла содержала два типа чередующихся морфологий чешуек — щетинкообразные и узкие, раздвоенные чешуйки. Тогда как в области непрозрачного крыла морфология чешуек напоминала «типичную» пигментированную чешуйку бабочки — плоская и яйцевидная с зазубринами на концах (1K и 1L).
Средняя (±s.d.) плотность чешуек на крыле взрослой особи была значительно ниже в обнаженной области (98.2±18.1 чешуек на мм2 у самцов и 102.3±17.2 у самок) по сравнению с непрозрачной областью (374.3±22.2 у самцов и 358.1±19.6 у самок) (1N).
В обнаженной области раздвоенные чешуйки были значительно меньше (498±39 мкм2) по сравнению со щетинкообразными чешуйками (831±183 мкм2), а в непрозрачной области чешуйки были самыми крупными (3467±382 мкм2) (1О).
Площадь обнаженной мембраны значительно различалась между областями крыла: 81.6±2.7 % у самцов и 82.2±4.3 % у самок в прозрачной области; 2.6±1.1 % у самцов и 1.4±0.7 % у самок в непрозрачной области (1P).
Изображение №2
Для изучения процессов развития крыльев и чешуек была проведена диссекция куколок Greta oto на разных этапах ее развития.
Как и у других видов чешуекрылых, развивающееся крыло Greta oto состояло из тонкого двойного слоя однородной эпителиальной ткани, и к 16 часам после формирования куколки (APF от after pupal case formation) многочисленные эпидермальные клетки дифференцировались с образованием параллельных рядов клеток-предшественников сенсорных органов (SOP от sensory organ precursor).
На данной стадии развития крыла было замечено, что прозрачная область крыла содержала более низкую плотность SOP клеток по сравнению с непрозрачной областью (2B и 2C). На площади 400 мкм2 плотность SOP клеток в прозрачной области составила 65.2±7.0, а в непрозрачной области — 169.2±15.7.
Из этого наблюдения следует, что на ранних стадиях развития крыла формирование паттерна SOP клеток по-разному регулируется между прозрачными и непрозрачными областями, что влияет на плотность чешуек взрослых особей и на площадь мембраны, обнаженной в разных частях крыла.
Далее были исследованы клеточная и цитоскелетная организация во время роста чешуек в прозрачных и непрозрачных областях крыла (2D-2I).
Было выявлено, что общие аспекты развития чешуек у Greta oto аналогичны тем, что обнаруживаются у разных видов бабочек и мотыльков, с некоторыми заметными различиями в росте чешуек именно в обнаженных областях крыла. На 30 часу APF клетки SOP разделились с образованием клеток чешуек и розеток* (2D и 2E). Клетка чешуйки находится внутри крыла, в то время как клетка впадины, связанная с каждой клеткой чешуйки, находится на поверхности.
Розетка* — пограничная эпителиальная клетка, образующая дистальный колпачок на покрывающей клетке, чтобы связать оболочку сенсиллы (органы чувств беспозвоночных) с соседней гиподермой через адгезивные контакты.Окрашивание фаллоидином показало появление небольших цилиндрических выростов чешуи, содержащих филаменты F-актина, а WGA окрашивание показало очертания мембраны, когда выросты чешуи начинают выступать и удлиняться за пределы поверхности крыла.
На этой стадии чешуйки в прозрачной области крыла оказались морфологически сходными с неспециализированными непрозрачными чешуйками: удлиненные и шаровидные с многочисленными мелкими актиновыми стержнями, отходящими веером от ножки к апикальному концу чешуи (2D и 2E). На 48 часу выросты чешуек удлинились (2F и 2G).
Флуоресцентное окрашивание выявило более крупные пучки F-актина (микрофиламенты*) на передней стороне (обращенной к мембране крыла) чешуек по сравнению с нижней стороной (видео ниже).
Микрофиламенты* — нити, состоящие из молекул глобулярного белка актина и присутствующие в цитоплазме всех эукариотических клеток.На этом этапе чешуйки в разных частях крыла приобретали совершенно разную морфологию. Чешуйки в обнаженной области вытянулись в вертикальной ориентации и приобрели два типа чередующихся морфологий: короткие и треугольные или длинные и щетиновидные выросты (2Е). В непрозрачной области чешуйки приобрели округлую и уплощенную морфологию, при этом нижние чешуйки были короче покровных (2G). На 60 часу выросты чешуек стали еще больше (2H и 2I).
Треугольные чешуйки в прозрачной области крыла начали формировать две новые ветви, которые раздваивались и удлинялись на концах в обоих направлениях, в то время как щетиновидные чешуйки удлинялись и изгибались (2Н). В непрозрачной области чешуйки были длиннее, шире и более плоские, с зазубринами на концах (2I).
На следующем этапе исследования был проведен ультраструктурный анализ развивающихся щетинок, раздвоенных и непрозрачных чешуек. Для выявления ультраструктурных деталей морфологии с помощью ПЭМ (просвечивающий электронный микроскоп) рассматривалось крыло Greta oto через 48 часов APF (изображение №3).

Изображение №3
Щетинковидные чешуйки на прозрачных участках крыльев были круглыми в поперечном сечении (3А-3С). Микроскопия также позволила увидеть дистальные и базальные участки щетиновидных чешуек, последние из которых показали на поперечном срезе наличие розетки (3B и 3C).
В дистальных отделах щетиновидной чешуйки микрофиламенты были больше на нижней стороне* крыла по сравнению с верхней* (3B), в то время как у основания щетиновидных чешуек микрофиламенты были более равномерно распределены по периферии (3C).
Нижняя сторона (adwing side)* — слой в структуре, представляющий собой гладкую, относительно плоскую пластину, обращенную к подложке крыла.С помощью ПЭМ удалось обнаружить несколько типов чешуек, имеющих более треугольную форму. Вероятно, это именно те чешуйки, которые в последствии станут раздвоенными в прозрачной области крыла (3D и 3E). Они были окружены периферическими микрофиламентами, которых было больше на нижней стороне.
Верхняя сторона (abwing side)* — слой, состоящий из массива гребней, идущих параллельно продольной оси чешуйки и соединенных поперечными ребрами, что вместе образует двумерную решетку.
Если же говорить про непрозрачные чешуйки, то их идентифицировать было достаточно просто ввиду больших габаритов и уплощенной морфологии (3F и 3G). В отличие от щетиновидных чешуек, крупные уплощенные непрозрачные чешуйки, по-видимому, содержат популяции микротрубочек, которые были более широко распределены и менее плотны.
Во всех типах чешуек наблюдалось наличие гексагонально упакованных F-актиновых филаментов и многочисленных внутренних органелл и везикул, включая митохондрии, электронно-плотные везикулы и свободные рибосомы.
Далее ученые рассмотрели онтогенез наноструктур мембран крыльев. Прозрачные области крыльев Greta oto содержат наностолбики, покрывающие поверхность мембраны (1I).

Изображение №4
Чтобы получить представление о развитии этих наноструктур, ученые исследовали поверхность эпителиальных клеток мембраны крыла с помощью ПЭМ микроскопии (4B-4F).
На 60 часу APF на перпендикулярном срезе эпителия крыльев была обнаружена непрерывная эпителиальная пластинка (4В и 4С). Эпителиальные клетки содержали микроворсинки, которые выглядели как тонкие линейные расширения от внутренних краев развивающихся клеток (4В и 4С). Поверхностный слой эпителия представлял собой внеклеточную ламеллярную систему, а выпячивания пластинок проявлялись на срезе в виде куполов дистальнее микроворсинчатых отростков (4С).
На 72 часу APF наблюдался тонкий наружный слой эпикутикулы, который возвышался над эпидермальными клетками, а на 120 часу было обнаружено, что этот слой над микроворсинками содержит куполообразные выпячивания и утолщенную кутикулу, возможно, секретирующуюся из равномерно расположенных микроворсинок (4D и 4E).
Наконец, на поперечном срезе полностью развитого взрослого крыла Greta oto, полученном с помощью ПЭМ, было обнаружено, что поверхность мембраны содержит куполообразные нановыступы с морфологией, сходной с морфологией массивов нановыступов на поверхности роговицы глаза насекомых (далее « сосковидные наноструктуры»), которые отличались более неравномерным распределением по высоте (4F).

Изображение №5
Основываясь на результатах электронной микроскопии мембранных наноструктур, ученые приступили к рассмотрению топографической организации и биохимического состава поверхности крыла взрослой особи Greta oto.
Для этого необходимо было обработать крылья двумя способами:
- физически удалить наноструктуры с поверхности, осторожно нажимая и растирая крыло между бумагой и пенополистиролом;
- использовать органические растворители, включая гексан и хлороформ, для извлечения липидов.
СЭМ (сканирующий электронный микроскоп) подтвердил, что первый метод частично или полностью удалил наноструктуры на поверхности мембраны крыла (5В). В области частичного удаления идентифицировались более мелкие куполообразные наноструктуры под верхним слоем наностолбиков (5Bi).
СЭМ химически обработанной поверхности крыла показала, что верхний слой наностолбиков неравномерного размера был полностью удален, обнажая слой равномерного расположенных куполообразных наноструктур, которые не растворялись при воздействии хлороформа или гексана (5C и 5Ci).
Из этого наблюдения можно сделать вывод, что верхний слой наностолбиков неравномерного размера состоит из секретируемого материала на основе воска, который расположен над меньшими наноструктурами на основе хитина.
Дабы проверить эту гипотезу, ученые экстрагировали поверхностный слой прозрачных областей крыла Greta oto гексаном или хлороформом, а затем проанализировали химический состав с помощью ГХ-МС (газовая хромато-масс-спектрометрия).
Химический профиль, созданный гексаном и хлороформом давал аналогичные результаты (5D). Во всех экстрактах можно было идентифицировать два алкана с прямой цепью, которые составляют примерно две трети обнаруженных соединений: 41.64±5.75% пентакозана (C25H52) и 23.32±5.35% гептакозана (C27H56). Остальные соединения в основном состоят из немного более крупных метилразветвленных алканов (монометил и диметил C27, C29 и C31) и сложных эфиров.
Следовательно, у Greta oto имеется два компонента ультраструктуры поверхности крыла: сосковидные наноструктуры на основе прокутикулы и верхний эпикутикулярный слой наностолбиков неравномерного размера, состоящий в основном из алканов с прямой цепью (5D и 5E).

Изображение №6
Чтобы выяснить, играют ли наностолбики на восковой основе роль в отражательной способности крыльев, ученые измерили спектры отражения необработанных и обработанных гексаном крыльев (изображение №6). Дополнительно была измерена геометрия наноструктур и толщина мембраны в поперечном сечении, чтобы определить среднее расстояния между двумя соседствующими наноструктурами, их высоту, радиус и т.д. (6B и 6D).
На основе СЭМ-снимков обработанных и необработанных образцов было смоделировано три архитектуры крыла, состоящие из:
- наностолбиков переменной высоты вместе с сосковидными наноструктурами на мембране крыла;
- сосковидных наноструктур на мембране крыла;
- мембрана крыла без каких-либо наноструктур.
Это позволило смоделировать различные оптические свойства крыла (6Е). Данные моделирования отражательной способности необработанных и обработанных образцов были очень схожи с экспериментальными.
В необработанных крыльях было обнаружено, что прозрачные области имеют низкое общее диффузное отражение, составляющее примерно 2%, что согласуется с предыдущими измерениями коэффициента отражения этого вида (6F). Напротив, обработанные гексаном крылья без верхнего слоя восковых наностолбиков имели примерно в 2.5 раза большую отражательную способность по сравнению с необработанными крыльями (6D и 6F). Из этого следует, что нерегулярная архитектура мембраны крыла и неравномерные восковые наностолбики на поверхности крыла Greta oto значительно улучшают антиотражающие свойства.

Изображение №7
Дополнительно были проведены аналогичные тесты, но с крыльями других видов бабочек: Godyris duilia, Haetera piera, Chorinea faunus и Hemaris thysbe.
У G. duilia и H. piera прозрачная поверхность мембраны крыла покрыта неравномерно расположенными наностолбиками (7A и 7B). После обработки гексаном крылья стали более отражающими, верхний слой неравномерно расположенных наностолбиков был удален, в то время как сосковидные структуры остались. Это подтверждает, что наностолбики основаны на воске, как и у Greta oto.
А вот у C. faunus и H. thysbe после обработки гексаном отражательная способность крыльев и регулярно расположенные наноструктуры никак не изменились. Следовательно, эти наноструктуры основаны на хитине (7C и 7D).
В обоих вариантах достигается прозрачность крыла, несмотря на отличные механизмы ее достижения. Это говорит о том, что подобные особенности возникли в филогенетически отдаленных линиях чешуекрылых.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Также можете ознакомится с исследованием, в котором проводилось сравнение разных видов бабочек, обладающих частично и полностью прозрачными крыльями.
Эпилог
В данном труде ученые попытались выяснить секрет прозрачности крыльев некоторых видов бабочек. Крылья чешуекрылых, как намекает нам само название этого отряда насекомых, содержат микроскопические чешуйки, которые и дают им столь разнообразный окрас. Однако у некоторых видов эти чешуйки обладают измененной морфологией (формой, размерами и т.д.). В добавок к этому прозрачные участки крыльев покрыты дополнительным слоем, состоящем из воска или хитина (в зависимости от вида), что еще больше усиливает антиотражательную способность крыла.
Факт того, что у разных видов прозрачнокрылых бабочек реализованы разные механизмы достижения прозрачности крайне любопытен. Это говорит о том, что когда-то в ходе эволюции образовалось две ветви бабочек, которые решили параллельно реализовать одну и ту же тактику камуфляжа, но реализовать по-своему. В будущем ученые намерены более детально рассмотреть именно этот эволюционный аспект столь необычной способности.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Комментарии (2)

mctMaks
04.02.2022 14:05а есть отличия от крыльев тех же стрекоз/мух/пчел? у них крылья тоже в достаточной степент прозрачны.
leonidru
Так выяснили или нет? Если да то в чем же секрет?