

Бескрайние просторы Вселенной таят множество секретов, которые нам, возможно, никогда не удастся раскрыть. Но что нам далекий Космос, если даже на родной планете, где мы обитаем уже не первую тысячу лет, есть места, покрытые завесой таинственности. И чем меньше мы знаем, тем сильнее наш страх. Как говорится, мы не боимся темноты, мы боимся того, что может в ней скрываться. Вполне вероятно мы исследуем и изучаем не из-за любопытства, праздного или здравого, а из-за страха, переполняющего нас, когда мы смотрим в кажущуюся бесконечной бездну. Как тут не упомянуть океан, а точнее его глубины. Жизнь на дне, где нет света, почти нет еды, где сама жизнь, будучи изощренно прекрасной, с каждым новым ответом порождает десяток новых вопросов. Группа ученых из Род-Айлендского университета (США) провела исследование бездны, выявив организмы, способные существовать на глубине 1200 м и при температуре 120 °С. Что это за организмы, и как им удается выживать в столь суровых условиях? Ответы на эти вопросы мы найдем в докладе ученых. Погружаемся.
Основа исследования
Мировая литературная классика полнится произведениями, так или иначе связанными с океаном и его глубинами. В каждом из произведений океан предстает чем-то могучим и непреодолимым, таинственным и манящим, завораживающим и пугающим. При упоминании океана сразу всплывают «Моби Дик» Германа Мелвилла, «Старик и море» Эрнеста Хемингуэя и многочисленные творения королей ужаса Говарда Лавкрафта и Эдгара Аллана По.

Детище Лавкрафта, не нуждающееся в представлении.
Столь трепетное отношение к океану вполне объяснимо. Он всегда рядом, он был до нас и, вполне возможно, будет и после. Ученые прикладывали максимум усилий, чтобы разгадать его тайны, на протяжении веков.

Водолазный колокол Сполдинга (иллюстрация из журнала «The Saturday Magazine», 1839 год), прародитель которого был впервые описанный Аристотелем еще в IV веке до н.э.
В наши дни изучение глубин стало намного проще, ввиду технологического развития. Множество устройств, инструментов и оборудования было создано исключительно с целью глубоководных исследований. Естественно, одним из основных центров внимания подобных исследований являются организмы, живущие в этой среде.
Глубоководное бурение океана позволило
Как мы знаем, на дне океана имеются гидротермальные источники, вокруг которых обитают организмы, способные выдерживать температуру до 122 °C. Метаболизм этих экстремофилов поддерживается за счет потоков окислителей и восстановителей из самого источника. Но глубоко под дном таких источников живительной энергии попросту нет. Как отмечают сами ученые, даже в нефтяных резервуарах, богатых органическими веществами, микробная активность сходит на нет при температуре ~ 80 °C.
Чтобы понять, как микроорганизмы способны выживать на глубине 1200 м при 120 °C без каких-либо дополнительных источников энергии, ученые изучили донные отложения в Нанкайском желобе у мыса Мурото (Япония).

Нанкайский желоб (Nankai Trough).
Гемипелагические аргиллиты и туфы возрастом 16 миллионов лет и толщиной 600 м за последние 0.4 миллиона лет засыпались отложениями, образовавшими поверх них слой той же толщины. Первые признаки микробной жизни в теплых отложениях на глубине ~ 800 м и температуре от ~ 80 до 90 °C были обнаружены еще два десятка лет тому назад, однако ограниченность возможностей оборудования того времени не позволила изучить этот регион более детально. К счастью, оборудование за эти годы стало намного лучше.
Ученые организовали экспедицию при поддержке IODP (Международная программа изучения океана) в области C0023 (32°22.0018'N, 134°57.9844'E, глубина воды 4776 м), расположенной рядом с местом первого открытия микробной жизни в почве. Во врем бурения была достигнута глубина 1177 м ниже дна, где была зафиксирована температура 120 ± 3 °C. Производился забор образцов, которые в дальнейшем анализировались для получения данных о числе живых клеток.
Результаты исследования
В исследуемой зоне C0023 глубинный профиль концентраций клеток заметно отличается от глобальной тенденции постепенного уменьшения концентраций клеток, наблюдаемой в аналогичных глубоких, но существенно более холодных (< 30 °C) отложениях.
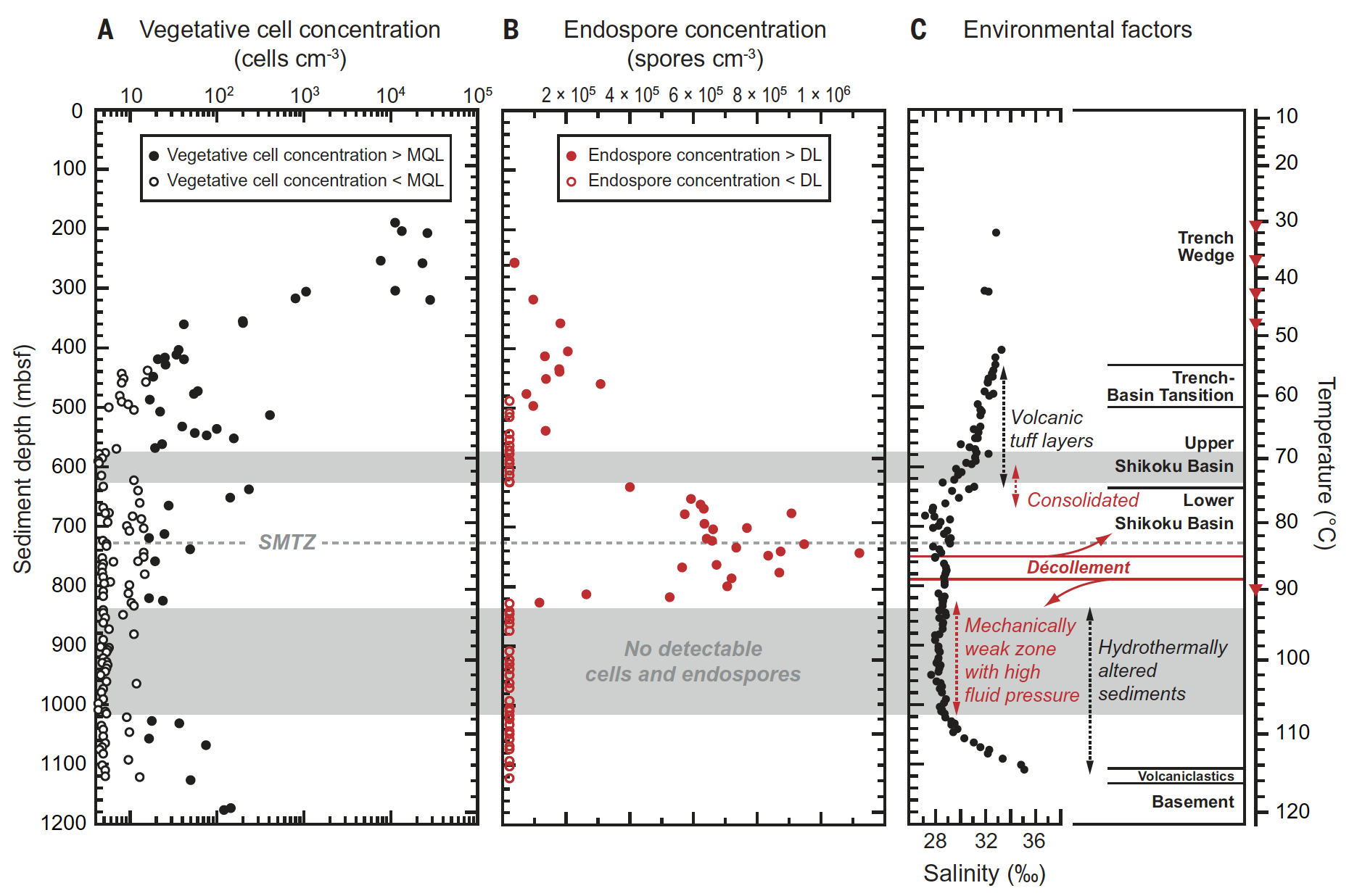
Изображение №1
На глубине ~ 300–400 mbsf (meters below sea floor, т.е. метров ниже морского дна) концентрации вегетативных клеток резко падают на два порядка и приближаются к MQL (minimum quantification limit, т.е. минимальный предел количественной оценки) при повышении температуры с 40 до 50 °C (1А).
Одновременно с этим концентрация эндоспор, т.е. спящих устойчивых структур, связанных с бактериальным типом Firmicutes, которые широко встречаются в морских отложениях и почвах, увеличивается до 2х105 клеток/см3 (1В). Тем не менее небольшая популяция микробов сохраняется при> 50 ° C в форме вегетативных клеток и эндоспор.
Любопытно, что вплоть до самого горячего (120 °C) слоя отложения, содержащие микробные сообщества с количеством вегетативных клеток до 400 клеток/см3, чередуются со слоями толщиной до 192 м, в которых клетки не были обнаружены вообще (1А).
Ученые отмечают, что подобные наблюдения хоть и наталкивают на вероятность загрязнения в ходе бурения, однако дополнительные исследования показали, что все обнаруженные клетки происходили именно из отложений (т.е. не были случайно занесены из другой среды).
В соответствии с чрезвычайно низкой концентрацией вегетативных клеток и сложностью извлечения ДНК из эндоспор, полученных ДНК было недостаточно для получения надежных данных о микробном сообществе, обитающем на глубине более 320 mbsf. В образцах с глубины менее 320 mbsf микробиом напоминал сообщества, обнаруженные в неглубоких подземных отложениях.
В отличие от рассеянного распределения вегетативных клеток в отложениях при температуре > 50 °C, эндоспоры демонстрируют четкую зональность (1B), что было количественно определено путем обнаружения биомаркера дипиколиновой кислоты (DPA). Вполне вероятно, что значительные уровни DPA могли накопиться после распада эндоспор, учитывая склонность 2-карбоксилированных пиридинов к декарбоксилированию при умеренном кратковременном нагревании.
Концентрация эндоспор заметно возрастает в слое отложений ~ 200 м с температурным интервалом от 75 до 90 °C, а максимум в 1.2х106 эндоспор/см3 наблюдался при 85 °C.
Среднее отношение эндоспор к вегетативным клеткам превышает 6000 в отложениях ниже 350 mbsf и, следовательно, на два-три порядка выше, чем в холодных донных отложениях. Вероятно, накопление эндоспор в отложениях, которые почти лишены вегетативных клеток, связано с термической историей исследуемого региона начиная с момента образования верхнего слоя отложений порядка 0.4 миллиона лет назад. При этом анализ породы показал, что возраст микробной активности составляет порядка 16 миллионов лет. Еще один любопытный факт заключается в том, что в слоях на глубинах от 570 до 633 mbsf и от 829 до 1021 mbsf не было обнаружено ни вегетативных клеток, ни эндоспор.

Изображение №2
Высокая концентрация метана со средним изотопным составом углерода (?13C-CH4) в -61.3 ± 3.0 ‰ (2A и 2B) указывают на биогенный метаногенез, по крайней мере, до температуры от 80 до 85 °C на глубине ~ 730 mbsf (SMTZ, т.е. до зоны перехода сульфат-метан).
Положительное изменение ?13C-CH4 в SMTZ (2B) указывает на биогенный сток метана и согласуется с предыдущими наблюдениями, которые продемонстрировали активность термофильных анаэробных окисляющих метан сообществ при этих температурах.
Ниже зоны SMTZ метан присутствует только в микромолярных концентрациях, с увеличением значений ?13C-CH4 и уменьшением соотношений метан/этан, указывающих на относительное увеличение термогенных углеводородов (2B).
В исследуемой области C0023 эксперименты с радиоактивными индикаторами выявили современную метаногенную активность в 65% исследованных образцов (2D). Потенциальные скорости метаногенеза за счет снижения содержания СО2 в отложениях на глубине ниже 300 mbsf, была ниже 4 пмоль/см3 в день. Эти данные соответствуют предыдущим исследованиям глубоководного дна.
Показатели метаногенеза варьируются в зависимости от исследуемой глубины. Самый низкий наблюдается ниже зоны SMTZ (< 0.6 пмоль). Если же опустится еще ниже, где нет следов клеток или эндоспор (2D), то метаногенез практически невозможно определить. Однако метаногенез вновь проявляет себя в трех самых глубоких зонах бурения, что дополнительно подтверждает существование активных метаногенных сообществ в отложениях при 110–120 °C.
Основным микробным субстратом в данном случае является ацетат (соли и эфиры уксусной кислоты). Ученые предполагают, что его образование из осадочного органического вещества при нагревании способствует развитию микробной жизни глубоко в донных отложениях океана.
При анализе образцов из разной глубины было выявлено, что концентрации ацетата и его углеродного изотопного варианта (?13C-ацетат) (2C) указывают на явные изменения в использовании ацетата с температурой и глубиной.
В слоях до 600 mbsf при температуре до 60 °C концентрация ацетата была достаточно низка (26 ± 22 мкМ). Это согласуется с устойчивым состоянием, управляемым микробиологическим производством и потреблением, как это наблюдается в других осадочных средах. Флуктуации ?13C-ацетата при среднем значении в -25.5 ± 3.4 ‰ указывают на продолжающуюся метаболическую активность. В слоях, где температура от 60 до 100 °C, использование ацетата минимально. Однако в температурном диапазоне 60…75 °C концентрация ацетата резко возрастает с одновременным снижением концентрации метана и накоплением эндоспор. Это позволяет предположить, что потребление микробов больше не уравновешивает высвобождение ацетата из осадочного органического вещества.
Ниже зоны SMTZ уровень концентрации ацетата составляет 9.2 ± 2.4 мкМ с неизменным значением ?13C-ацетата около -18.8 ± 0.5. Комбинация высокой концентрации и низкой изотопной изменчивости подразумевает наличие пула ацетата без существенного оборота в зоне с преобладанием эндоспор, а также в нижележащей зоне толщиной 200 м, где не было обнаружено ни клеток, ни эндоспор.
Однако ниже 1030 mbsf концентрации ацетата снижаются, а ?13C-ацетат линейно увеличивается с глубиной, достигая максимума в -7.9 ‰ в самой глубокой пробе поровой воды, извлеченной из 1101 mbsf. Эта тенденция согласуется с активными гипертермофилами, разлагающими преимущественно 13C-обедненный ацетат, оставляя остаточный ацетат изотопно обогащенным. Без постоянного потребления диффузия гомогенизировала бы вариации ?13C-ацетата, как это наблюдается в вышележащих отложениях.
Учитывая клеточные концентрации от 10 до 100 клеток/см3 в отложениях, требуемые скорости клеточного метаболизма на два-три порядка ниже, чем наблюдаемые в лабораторных культурах гипертермофильных архей Pyrococcus furiosus и Archaeoglobus fulgidus, но на два-три порядка выше, чем скорости в глубоких отложениях с температурами < 30 °С.
Из этого следует, что существует небольшое микробное сообщество, использующее ацетат при температуре > 100 °C. При такой температуре микробы, вероятно, требуют больше энергии, а потому перерабатывают субстрат быстрее, чем при более низкой температуре.
Вышеописанные результаты дают понять, что существует тесная связь между микробной жизнью, температурой и глубиной ниже уровня дна. Примером этого является массовый коллапс популяции вегетативных клеток в отложениях возрастом < 0.4 млн. лет на глубине от 300 до 400 mbsf. Этот диапазон глубин (температура от 40 до 50 °C) находится в пределах верхнего диапазона роста мезофилов*.
Мезофилы* — организмы, лучше всего растущие при умеренной температуре (от 20 до 45 °C).Повышение концентрации эндоспор в результате споруляции мезофильных эндоспорообразующих Firmicutes подтверждает вывод о том, что численность микробных популяций в основном контролируется зависимыми от температуры физиологическими факторами вплоть до глубины в 600 mbsf. Глубже этого уровня дополнительные эффекты могут оказывать геологические факторы.
Резкое снижение биогенного метаногенеза и утилизации ацетата при температуре от 70 до 75 °C совпадает с верхней температурной границей роста термофилов*. Любопытно то, что этот интервал глубин одновременно охватывает литологическую границу между Верхним и Нижним бассейнами острова Сикоку.
Термофилы* — организмы, способные существовать при постоянно высоких температурах (выше 45 °C).На этом литологическом стыке туфы (затвердевший вулканический пепел) более не присутствуют. При изменении туфа образуется смектит, а микробное восстановление Fe(III) в смектите служит энергоемким процессом, способствующим превращению смектита в иллит на глубине от 500 до 600 mbsf. Следовательно, изменения некоторых микробных сообществ могут быть обусловлены микробно-минеральными взаимодействиями.
В верхнем слое отложений толщиной 200 метров при температуре от 90 до 100 °C была выявлена зона без клеток или каких-либо геохимических признаков микробной активности. В этой зоне недостаточно уплотненные и механически слабые отложения подвергаются избыточному давлению и кратковременному воздействию горячих потоков от ~ 145 до 220 °C. Периодическое нагревание может создавать стерильные участки отложений, однако микробные клетки, потребление ацетата и метаногенная активность проявляются в отложениях с температурой > 100 °C, где механическая прочность и соленость возрастают по направлению к поверхности интерфейса донных отложений.
Гидравлическое сообщение между базальтами и вышележащими отложениями подтверждается наличием эпигенетической минерализации в виде жил кальцита и железистых оксидов металлов. За счет этого имеется массоперенос между базальными отложениями и базальтовым водоносным горизонтом*.
Водоносный горизонт* — осадочная горная порода, представленная одним или несколькими слоями горных пород с различной степенью водопроницаемости.За счет этого снижается давление и происходит восполнение субстратов, таких как восстановленное железо и сульфат. Данный процесс и позволяет микроорганизмам развиваться в средах, которые в противном случае были бы необитаемы.
Для более детального рассмотрения нюансов исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня труде ученые поделились своими находками, полученными в ходе исследовательского бурения до глубины 1777 метров ниже дна океана. Как оказалось, даже там есть жизнь, что особенно удивительно, если учесть местный жаркий «климат», ибо температура в глубоких слоях отложений составляет порядка 120 °C.
На первый взгляд, в таких условиях не должно быть признаков жизни, однако ученые обнаружили наличие микроорганизмов, которые приспособились к суровой среде обитания. Но самое любопытное в том, что на меньших глубинах с температурой около 45 °C не было обнаружено никаких организмов. Другими словами, дно океана напоминает записную книжку эксцентричного писателя, решившего написать пару строк на первой станице, пропустить десяток, и продолжить с середины.
Основным двигателем жизни на глубине почти 1200 метров ниже дна при температуре 120 °C ученые считают гидродинамическую взаимосвязь между слоями отложений, которая позволяет наполнять более глубокие слои питательными веществами.
Авторы исследования не боятся открыто говорить о том, что полученные результаты являются лишь малой долей того, что можно будет открыть в будущем. Единственное препятствие на пути к этому — технологии. На данный момент пока еще нет инструментов, позволяющих полноценно проанализировать образцы, полученные в ходе глубоководного бурения. Слишком много данных остаются неучтенными, что влияет на понимание общей картины происходящего в столь чуждой для человека среде. Посему ученые намерены направить все свои усилия на решение этой проблемы.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


drWhy
«дно океана напоминает записную книжку эксцентричного писателя, решившего написать пару строк на первой станице, пропустить десяток, и продолжить с середины.»
Возможно, мы пока просто не находим следы жизнедеятельности живых организмов с пропущенных страниц. Ну и поаккуратнее нужно листать эту книжку — вполне возможно, что обитатели разных страниц не вполне совместимы.
В цикле фантастических романов «Рифтеры» Питер Уоттс, гидробиолог с докторской степенью, описывает результат заселения древнего микроогранизма на основе РНК, высвобожденного в результате строительства геотермальной станции на дне океана в месте разлома, в клетки человека, как некогда туда попали митохондрии. Человек, а именно слегка модифицированные глубоководные ныряльщики, обслуживающие станцию, стали идеальным передаточным звеном между древней жизнью, обитающей под океанским дном, и наземной экосистемой. В итоге более простая и эффективная форма жизни вытеснила более поздние и сложные.
Спасибо за предоставленную возможность ознакомиться с публикацией.
LeToan
Скорее будет наоборот, нежели уоттсовский вариант. Просто про реалистичный вариант фантастический остросюжетный сериал не напишешь.
drWhy
Полагаете, человечество угрожает существованию микроорганизмов на глубине километра под дном океана?
LeToan
При встрече — да.
micho1973
Ни в одном фантастическом произведении, в котором рассказывается о встрече с ранее вымершей, а потом внезапно возродившейся древней агрессивной и смертоносной жизнью, не объясняется, почему она проиграла в конкуренции с менее агрессивными и смертоносными формами и вымерла. Если она проиграла раньше, то с чего бы ей выиграть сейчас?
BigBeaver
Там ГМО вирус в рифтерах а не просто «занесли».
CherryPah
ну может ей метеорит на голову упал
а без этого фактора, она до сих пор бы доминировала
tendium
Ну… я вам приведу один пример: была такая большая группа животных как териодонты, которые были почти полностью вытеснены архозаврами (я не утверждаю, что они сыграли доминирующую роль, но тем не менее долгое время господствовали именно они). Но потом что-то пошло не так, и сравнительно небольшая линия, оставшаяся от ранее широко распространенных териодонтов, которую мы теперь знаем как млекопитающие, опять стала доминировать. А архозавры, хоть и не вымерли полностью (от них остались птицы и крокодилы), но теперь значительно менее разнообразны и явно уступили в доминировании.