

В августе 1913 года Петр Нестеров выполнил один из самых захватывающих, опасных и сложных маневров в авиации — мертвую петлю. На тот момент, особенно учитывая тогдашние технологии, да и сейчас это настоящий высший пилотаж. Многие современные профессионалы и любители из области авиации все бы отдали, чтобы побеседовать с Нестеровым. К сожалению, это невозможно, но с нами по соседству живут и частенько нам надоедают другие асы высшего пилотажа. Как правило, мы их либо не замечаем, либо гоняем по квартире с тапком или газетой, потому что для нас они вредители, переносчики всякой заразы и просто надоедливые летуны. Но для ученых эти существа являются хранителями секретов полета и посадки. Сегодня мы с вами познакомимся с удивительным исследованием умения обыкновенные мух приземляться вверх ногами на любой поверхности. Какой механизм посадки используют мухи, из каких процессов он состоит и насколько сложно его искусственно реализовать в робототехнике? Об этом мы узнаем из доклада исследовательской группы. Поехали.
Основа исследования
Конечно, нельзя отдавать все лавры первенства в летном деле мухам, поскольку многие другие насекомые также обладают своими уникальными способностями в сфере полета и посадки. Однако мухи так сказать аккумулировали в себе максимальное число таких умений, которые напрямую связаны с их анатомией.

Внешний вид мухи вида Calliphora vomitoria.
В данном исследовании главную роль сыграли мухи вида Calliphora vomitoria (синяя муха). Этот вид можно найти практически в любом уголке планеты: от Южной Америки до Восточной Азии. Габариты у этих мух довольно внушительные, 10-14 мм в длину, что примерно в два раза больше, чем у обычных комнатных мух (Musca domestica). Внешне отличить Calliphora vomitoria от ее сородичей можно по синему брюшку, благодаря которому она еще получила название «bluebottle fly», намекающее на сходство с цветом синего бутылочного стекла.
Помимо внешних отличий от комнатных мух у Calliphora vomitoria есть еще и поведенческие. Синие мухи предпочитают летать небольшим роем, нежели по одиночке, что позволяет им быстрее находить пищу (как правило это нектар сильно пахнущих цветов). Когда одна особь из роя находит пищу, она выделяет специальный феромон, сигнализирующий о находке всем остальным.
Люди часто используют личинки синих мух в качестве наживки во время рыбалки, но для насекомых это не самое уникальное и, по правде говоря, не самое достойное применение. Не только рыбаки находят пользу в синих мухах, но и криминалисты. Мухи, как мы знаем, не из брезгливых, потому трупы животных и людей их не пугают, а скорее привлекают. Для криминалистов это дополнительный критерий оценки времени смерти, так как синие мухи обладают очень четким взаимоотношением скорости роста их личинок и температуры окружающей среды. Зная температуру, можно по личинкам определить время, когда была сделана кладка яиц.
Но сегодня мы будем говорить не о криминалистике, а об авиации. Изучение инвертированной посадки, т.е. посадки на потолок, как ее называют ученые, проводилось и ранее, но не так детально. При этом все же были обнаружены некоторые любопытные нюансы. Во время наблюдений за инвертированной посадкой комнатных мух ученые заметили, что те совершают «полукруглые» вращательные движения перед контактом ипсилатеральных* лапок с поверхностью.
Ипсилатеральный* — расположенный на одной и той же стороне (утрировано, левая рука человека является ипсилатеральной по отношению к левой ноге).Следом за этим движением происходило латеральное колебание тела при помощи лапок, что приводило к соприкосновению остальных лапок с поверхностью. Эти и другие маневры перед фактической посадкой, естественно, обусловлены сенсорикой, т.е. зрительными, осязательными и другими сигналами, полученными мухой перед посадкой.
В рассматриваемом нами сегодня исследовании ученые из университета штата Пенсильвания провели ряд наблюдений за посадкой синих мух на инвертированной поверхности в полетной камере (в аэродинамической трубе) с использованием высокоскоростного видео (1А).
Изображение №1
Кинематика тела и крыльев мух была получена путем оцифровки анатомических меток (1B и 1C).
Результаты наблюдений
Прежде всего ученые решили вычленить и проанализировать поведенческие аспекты успешной посадки.
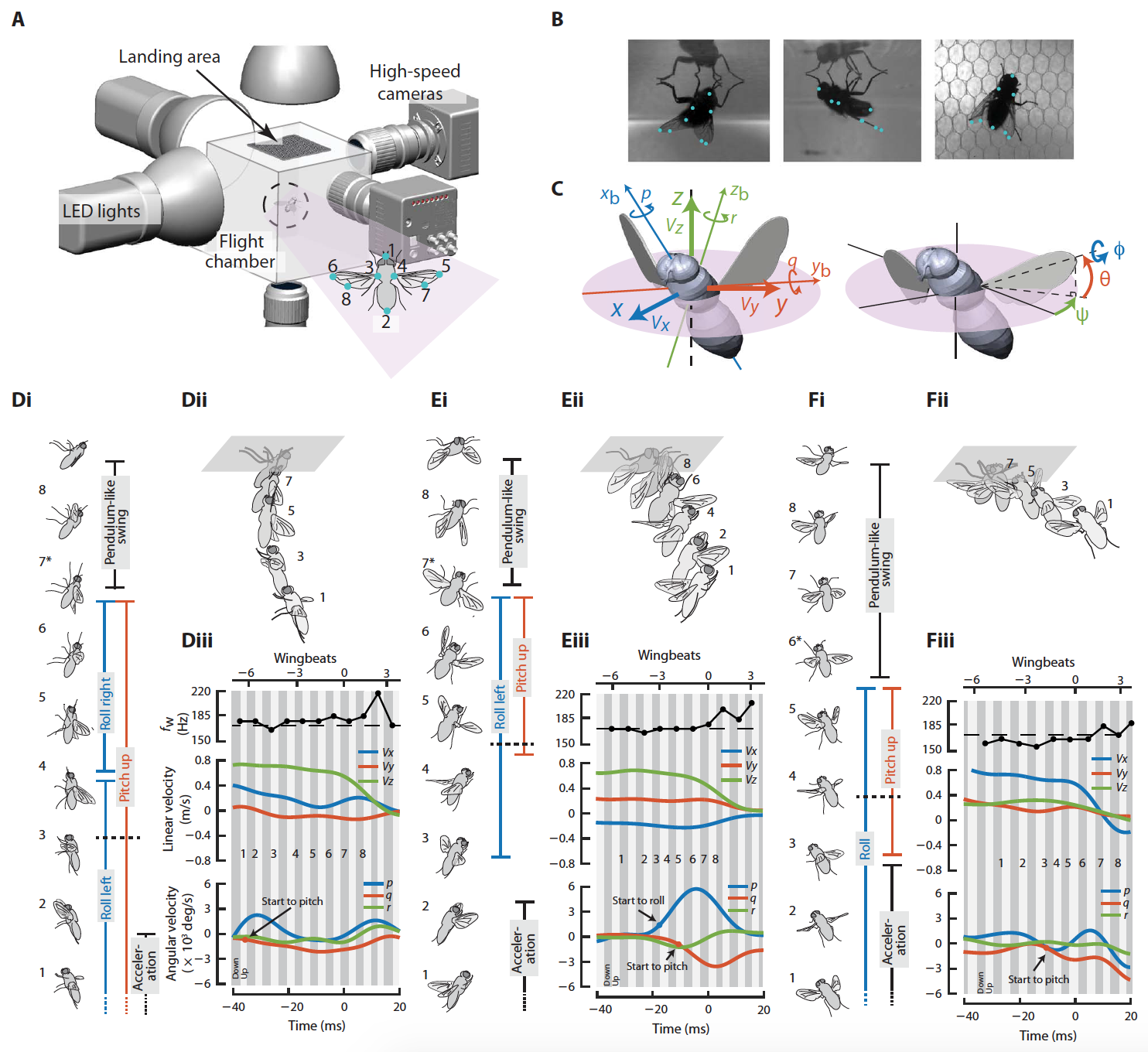
Они отмечают, что плодовые мушки приземляются на вертикальных поверхностях посредством непрерывного замедления движения и незначительного вращения тела перед посадкой. А вот синие мухи во время посадки демонстрировали сразу 4 последовательных поведенческих аспекта: вертикальное ускорение, вращательный маневр всего тела, разгибание лапок и покачивание всего тела при помощи передних лап, стабильно зафиксированных на поверхности. Длительность всего процесса успешной посадки составляла примерно 4-8 ударов крыла по данным 18 наблюдений, при этом средняя частота ударов крыла составляла 172.7 ± 7.7 Гц.
Во время успешных посадок ученые наблюдали также и существенные кинематические изменения осей вращения, величин вращательных маневров и степени раскачивания тела за счет лапок.
Собрав воедино все наблюдения, ученые выявили основные стратегии во время приземления, которые были классифицированы следующим образом: доминирующая по тангажу*, доминирующая по крену*, комбинированная и доминирующая по латеральному/продольному раскачиванию.
Тангаж* — угловое движение тела относительно горизонтальной поперечной оси.
Крен* — поворот тела вокруг его продольной оси.
Видео №1: приземление с преобладающим тангажом (соответствует изображению 1D).
Видео №2: приземление с преобладающим креном (соответствует изображению 1Е).
На изображениях 1D-1F показаны типичные модели поведения синей мухи во время посадки. На первых двух примерах мухи использовали быстрые вращательные маневры, главным образом, относительно оси тангажа (1D и видео №1) или крена (1Е и видео №2), что устанавливало их тело перед приземлением в почти инвертированную (брюшком кверху) ориентацию.
Выравнивание лапок происходило практически сразу после начала вращательных маневров (горизонтальные черные пунктирные линии на 1Di и 1Ei). После того, как лапки касались поверхности, начиналась фаза раскачивания тела.
Любопытно, что средняя угловая скорость вращательных маневров достигала приблизительно 4000 °/с, а максимальное ее значение могло превышать 6000 °/с, что значительно выше, чем у плодовых мушек или даже у колибри.
Видео №3: приземление с преобладающим продольным колебанием тела (соответствует изображению 1F).
В третьем варианте мухи почти полностью полагались на колебание тела при помощи лапок (1F и видео №3), а вращательный маневр был незначителен.
Видео №4: приземление с преобладающим латеральным колебанием тела.
Таким образом, по сравнению с первыми двумя вариантами, эта стратегия приземления, вероятно, использовала больше механических и структурных процессов (например, адгезия* из-за пульвилл* или демпфирование* из-за вязкоупругости лап), что способствовало передаче линейного импульса тела вращательному импульсу.
Адгезия* — сцепление поверхностей разных по происхождению (например, капли росы на траве).
Пульвиллы* — мягкие подушечки кончика лапок, каждая из которых расположена под своим коготком. Основная задача пульвилл это удержание насекомого на гладкой или наклонной, вертикальной поверхности.
Демпфирование* — искусственное подавление колебаний.Следующим шагом в исследовании стало изучение кинематических отличий между успешными приземлениями и неудачными.
Видео №5: приземление с комбинированной механикой крен-тангаж.
К неудачным приземлениям относились те, которые закончились столкновением с поверхностью (что для ученых данные для анализа, то для мух сотрясение мозга, образно говоря, конечно).
Изображение №2
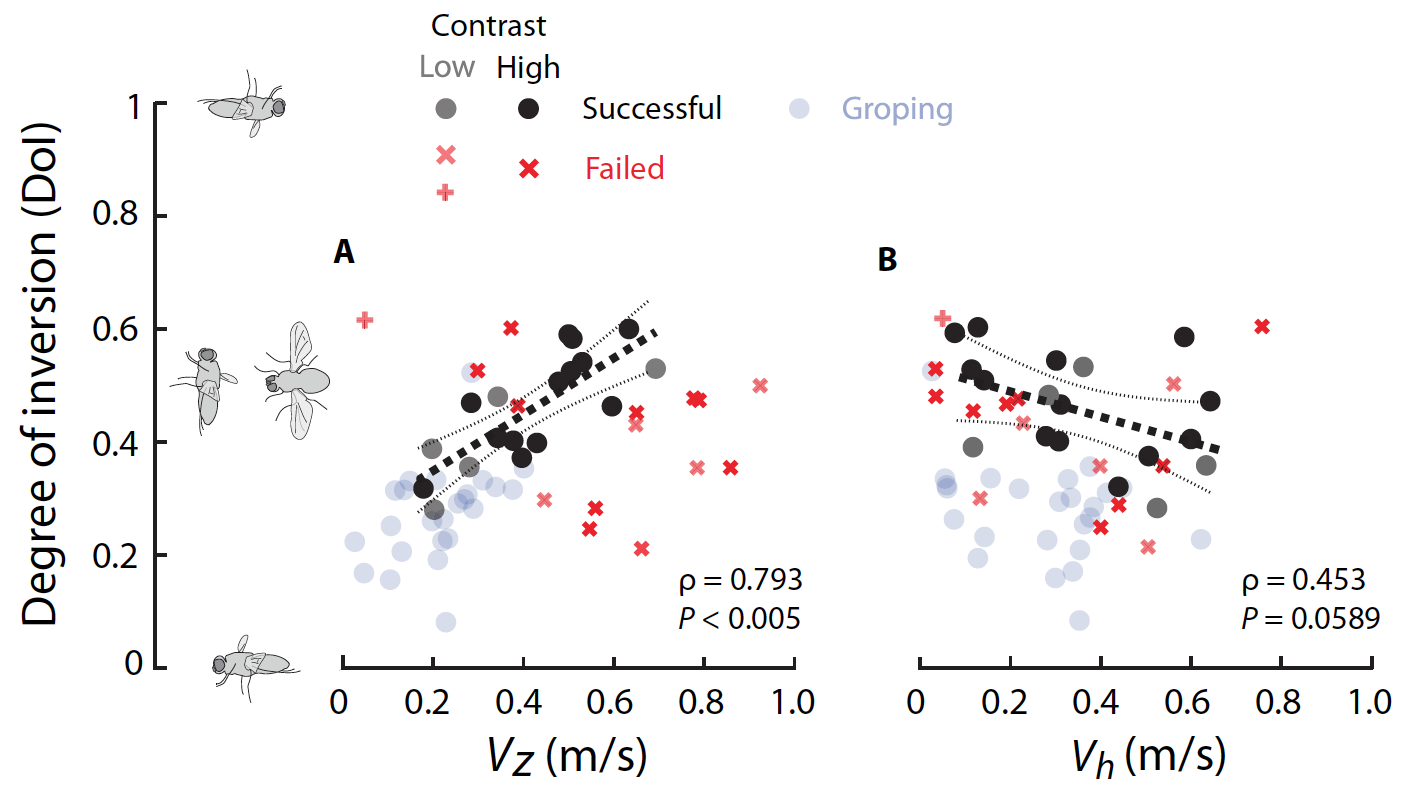
Для численного определения того, насколько хорошо было ориентировано тело мухи, была рассчитана степень инверсии (DoI от «degree of inversion»), измеряющая степень, в которой тело мухи выровнено относительно полностью инвертированной ориентации до приземления. DoI варьируется от 0, что означает отсутствие инверсии тела, т.е. брюшком вниз, до 1, что означает инверсию всего тела, т.е. брюшком вверх. При успешных посадках DoI увеличивался с вертикальной скоростью (2A), но уменьшался с продольной скоростью (2B).
Это наблюдение указывало на то, что мухам необходимо было больше переворачиваться, когда их восходящая скорость была выше или когда их горизонтальная скорость была ниже. В большинстве неудачных приземлений (всего их было 15) мухи были недостаточно перевернуты до момента приземления по сравнению с положением мух, совершивших успешные посадки.
Причиной неудач могло быть несвоевременное или недостаточное вращение тела перед посадкой. Таким образом, эти мухи не могли должным образом приземлиться на потолок, а врезались в него.
Видео №7: неудачное приземление из-за слишком раннего вращательного маневра.
Кроме того, неудачные посадки происходили еще и по причине слишком раннего вращения тела (видео №7), так как более ранняя инверсия приводила к недостаточной вертикальной скорости, необходимой для достижения потолка (отмечено знаком «+» на графиках 2А и 2В).
Видео №8: неудачное приземление из-за слабой инверсии тела с несвоевременным выравниванием лапок.
Еще одной причиной неудач, несмотря на правильную инверсию и скорость движения, было несвоевременное выравнивание лапок, что приводило к неправильному расположению и, следовательно, недостаточно цепкому контакту с поверхностью (видео №8).
Видео №6: пример контактного приземления на потолок.
Несмотря на все проколы и недочеты, некоторые мухи все же могли восстановиться и приземлиться успешно, используя контактную посадку. В таком случае мухи летят с низкой вертикальной скоростью под потолком, потом ощупывают поверхность, используя передние лапки, а потом цепляются и приземляются (видео №6).
Видео №9: неудачное приземление из-за слабой инверсии тела с незначительным вращением тела.
Суммируя данные, полученные из наблюдений, ученые пришли к выводу, что кинематические аспекты приземления мухи содержат в себе гораздо больше различных нейронных процессов, чем считалось ранее. Самой яркой особенностью по-прежнему остается вращательный маневр. Осталось только понять, что его провоцирует, а точнее какие сигналы приводят к тому, что муха применяет его для приземления.
Первыми на ум приходят, конечно, визуальные сигналы, которые мухи получают во время приближения к потолку с линейной скоростью, состоящей из трех компонентов (1С): вертикальная (Vz), продольная (Vx) и латеральная (Vy).
Изображение №3
Благодаря работам своих предшественников исследователи точно знают три основных типа визуальных подсказок, необходимых для оценки движения мухи.
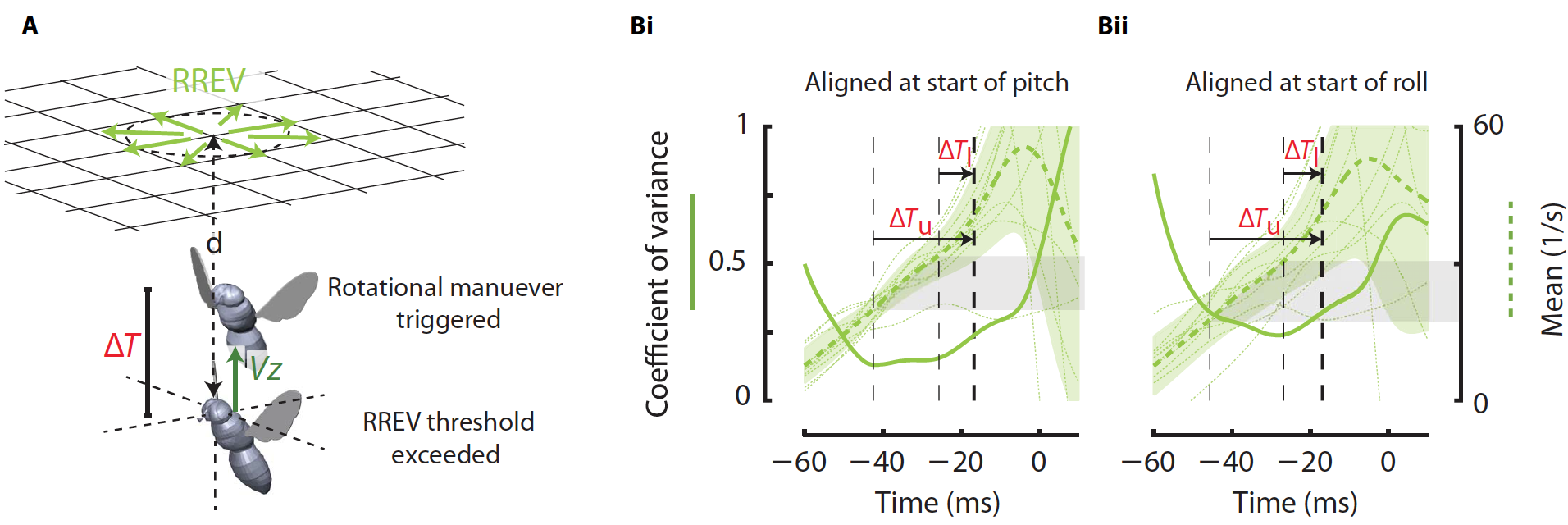
Во-первых, это относительная скорость расширения сетчатки (RREV) из-за неподвижных стимулов (3А). Этот показатель может быть рассчитан как отношение скорости увеличения цели к ее фактическому размеру на сетчатке мухи во время движения вверх (Vz). RREV также соответствует обратной величине времени до столкновения (t). RREV считается важным сигналом восприятия, который контролирует скорость приближения во время посадки или обхода препятствий.
Изображение №4
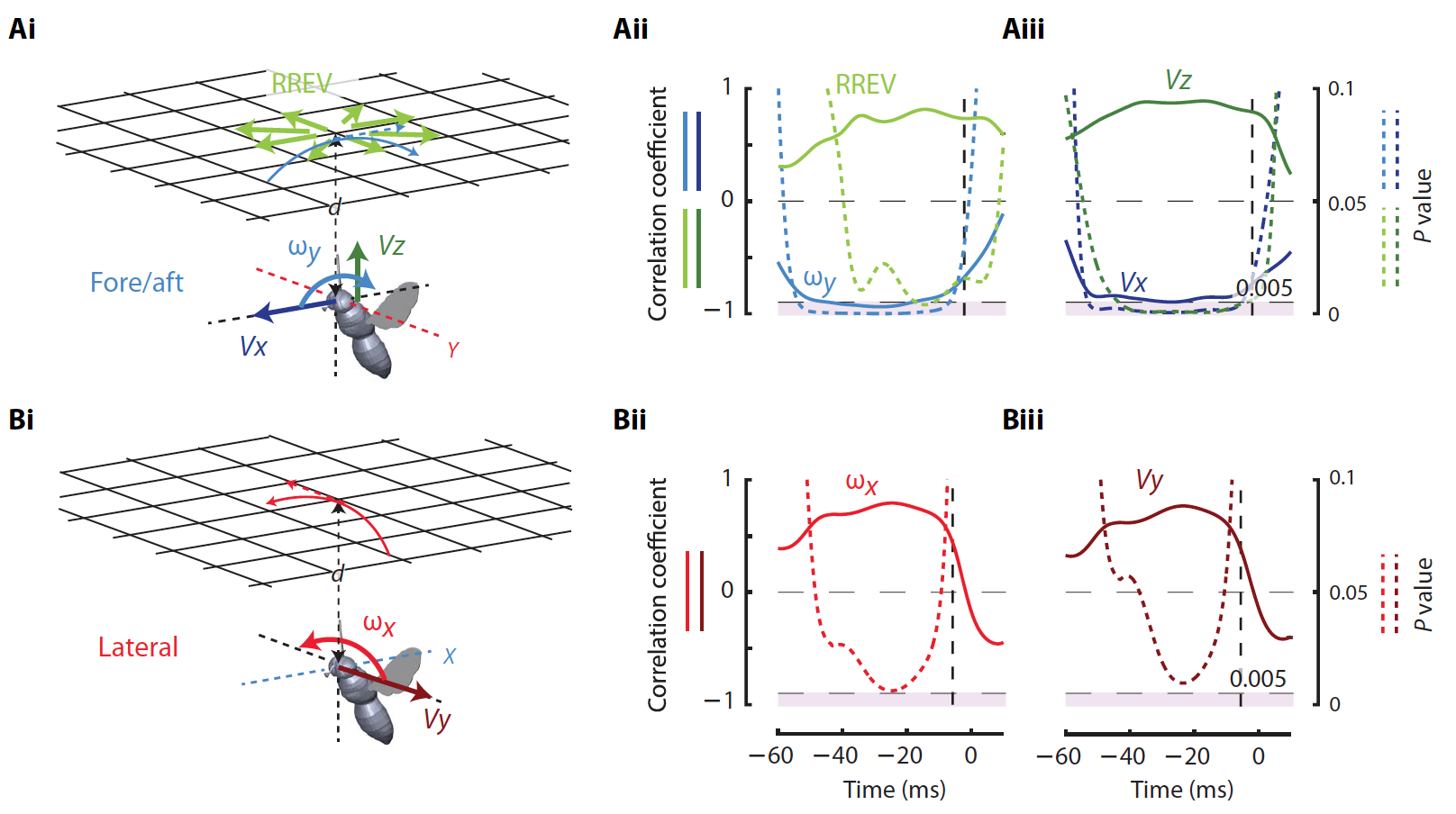
Во-вторых, относительная угловая скорость продольного направления (4Ai) потолка на сетчатке мухи (?y). Эти визуальные сигналы являются результатом перемещения тела вперед / назад (Vx), т.е. в горизонтальном направлении. Ранее было установлено, что именно эти сигналы регулируют приземление на горизонтальных поверхностях.
В-третьих, относительная поперечная угловая скорость потолка (?x) на сетчатке мухи (4Bi), полученная в результате латерального перемещения (Vy).
Исследователи предполагают, что вращательный маневр запускается после фиксированной временной задержки, когда провоцирующий (сигнал, что приводит к фактическому действию) визуальный сигнал достигает порогового значения. Такое предположение позволяет оценить взаимосвязь вышеописанных сигналов и маневров движения, выполняемых мухой во время посадки.
Первым делом были рассчитаны временные кривые коэффициента вариации (CV) каждого из визуальных сигналов. За основу были взяты наблюдения, во время которых мухи успешно приземлялись на потолке с первой попытки.
Если вращательные маневры были запущены после некоторой задержки, когда визуальный сигнал достиг порогового значения, CV этого визуального сигнала должен быть минимальным. Кроме того, он должен быть ниже, чем у других не провоцирующих визуальных сигналов, т.е. провоцирующий визуальный сигнал должен иметь наименьшую дисперсию. Наблюдения и расчеты показали, что среди трех протестированных сигналов именно RREV имел наименьшее значение CV в момент запуска поворота (15%) и крена (25%), а у двух других визуальных сигналов CV был значительно выше (3В).
Это означает, что RREV является основным визуальным сигналом, который провоцирует начало вращательного маневра во время инвертированного приземления.
Значение CV для RREV было минимальным в промежутке между 7 мс (?Tl) и 27 мс (?Tu) до начала вращательного маневра. Следовательно, визуальная задержка DT между моментом восприятия и началом маневра была в этом небольшом диапазоне. Также было установлено, что критическое время до столкновения, ниже которого запускается вращательный маневр (чтобы избежать столкновения и успешно приземлиться), составляет 31-53 мс. Для сравнения, у мух, приземляющихся на вертикальных поверхностях, этот показатель составляет 76 мс. Другими словами, при приземлении на потолок у мух значительно меньше времени на ошибку.
Вращательные маневры, судя по наблюдениям, были достаточно вариативными с точки зрения скорости крена и тангажа. Эта вариативность может говорить о том, что вращательный маневр регулируется не только временной составляющей, но и другими сенсорными сигналами.
Мухи используют сенсорные сигналы для обеспечения вращательного маневра через каналы прямой и обратной связи. Канал прямой связи выдает команды самопроизвольного управления, которые вызывают значительные переходные изменения движения крыла. Эти изменения напрямую влияют на амплитуду первичной реакции на маневр.
Канал обратной связи обеспечивает компенсационную реакцию, которая вызывает мелкие изменение движения крыла. Подобные изменения влияют на демпфирование (подавление колебаний) и на стабилизацию маневра.
Оценка степени влияния на маневр того или иного сенсорного сигнала была проведена путем выявления корреляции максимального крена и частоты вращения во время вращательного маневра с сенсорными сигналами, воспринимаемыми мухами до или после начала маневров.
К предлагаемым сенсорным сигналам относятся три визуальных (RREV, ?x и ?y), рассмотренных ранее, и три компонента линейной скорости (Vx, Vy и Vz).
Линейный регрессионный анализ показал, что в течение большого периода времени до начала маневров максимальная скорость тангажа была положительно коррелирована с RREV (4Aii) и вертикальной скоростью Vz (4Aiii), но отрицательно с продольным вращением ?y (4Aii) и продольной линейной скоростью Vx (4Aiii).
В свою очередь пиковые скорости крена положительно коррелировали с латеральным вращением ?x (4Bii) и латеральной линейной скоростью Vy (4Biii) во время продолжительного периода времени до и после начала маневра.
Проведенный анализ также указал на то, что мухи могут изменять силу вращательного маневра для того, чтобы эффективнее использовать колебания тела в момент посадки, т.е передачу линейного импульса угловому.
Как показали наблюдения, скорость тангажа снижалась, когда линейный импульс был высоким, на что указывает отрицательная корреляция между пиковым уровнем тангажа и сенсорными сигналами. Таким образом, вместо тангажа, мухи переворачивают свое тело, увеличивая продольное колебание тела при помощи лапок и передачу прямого линейного импульса к угловому импульсу тангажа.
Также наблюдалось повышение скорости крена, когда латеральный линейный импульс был высоким, на что указывает положительная корреляция между пиковой скоростью крена и сенсорными сигналами. Такая обратная корреляция может возникать в ситуациях, когда мухам необходимо большее расстояние, чтобы поднять их ипсилатеральные лапки достаточно высоко и достичь потолка для дальнейшего раскачивания тела.
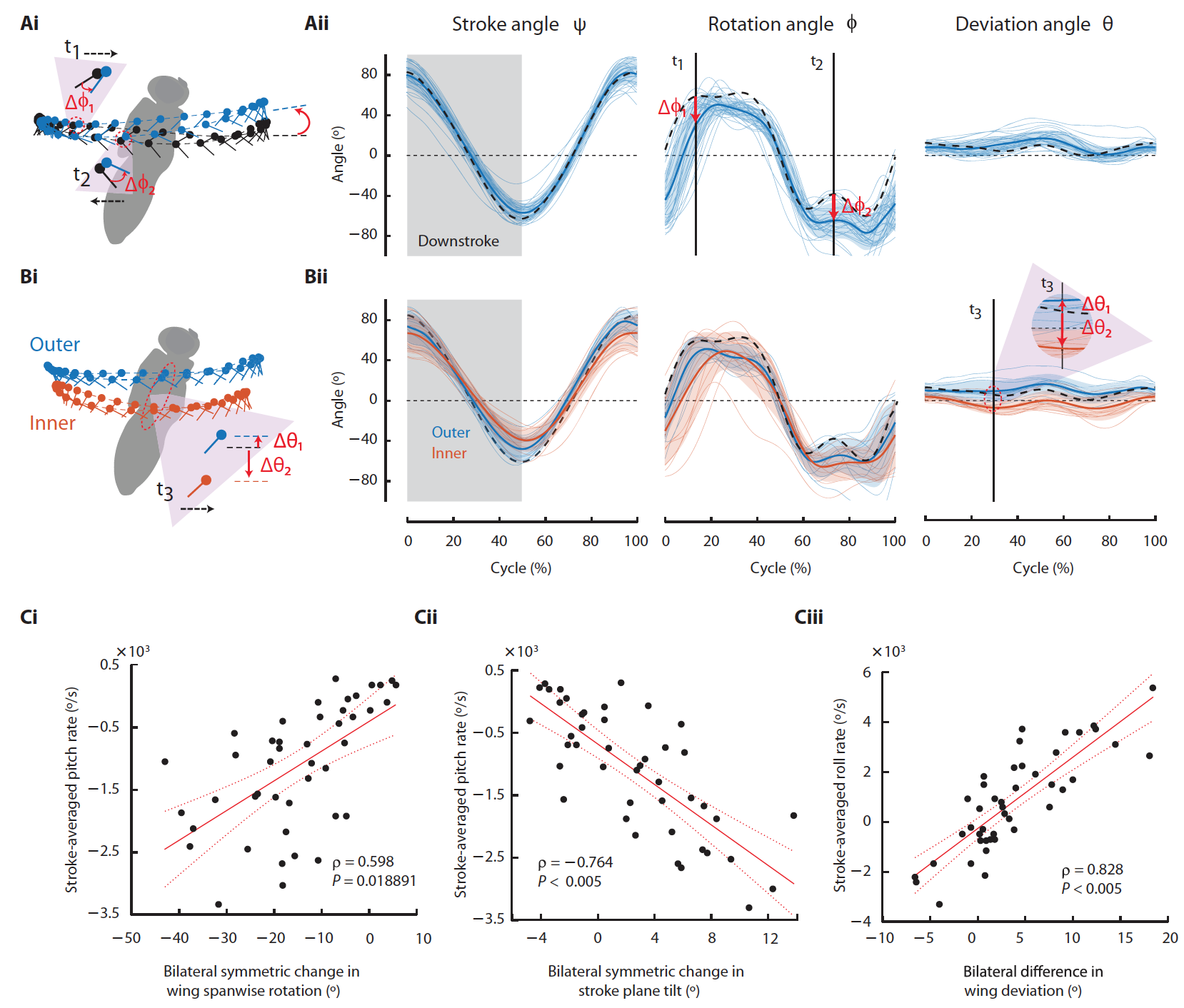
Не стоит забывать и про главнейший атрибут полетов для мух, т.е. про крылья. Исследователи проанализировали кинематику крыльев мух во время приземления на потолке, поскольку именно крылья создают различные аэродинамические импульсы, необходимые для успешных маневров.
Наблюдения за крыльями синих мух выявили ряд изменений, происходящих в момент маневра.
Изображение №5
Первое, что было обнаружено, это двустороннее симметричное изменение в продольном вращение крыла (5А и 5Сi) и двусторонние симметричные изменения угла плоскости взмаха (5А и 5Сii), которые оба сильно коррелировали с частотой тангажа. Также были изменения наклона крыла (5B и Ciii), которые сильно коррелировали со скоростью крена. Кроме этого, были еще и изменения в амплитуде взмахов и вращения среднего крыла, что способствует крену.
Изображение №6
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
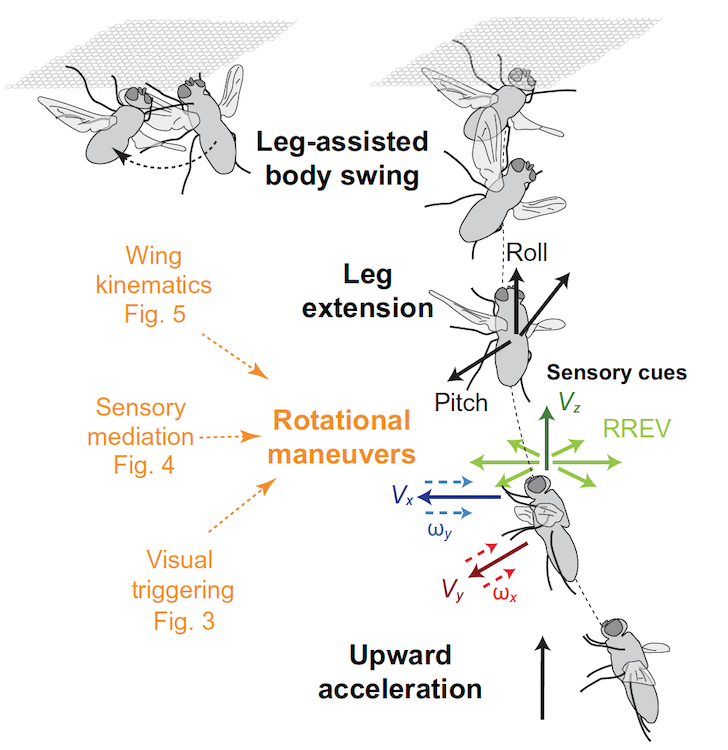
Суммируя все наблюдения и результаты расчетов, исследователи пришли к выводу, что инвертированное приземление на потолок происходит за счет хорошо скоординированной последовательности поведенческих модулей (схема выше).
Приземление на потолок может включать в себе нейронные процессы, которые не только вычисляют RREV, кодирующий информацию о времени до столкновения, но и другие сенсорные сигналы, которые предположительно кодируют многоосевое линейное перемещение тела.
На данный момент ученые пока не могут точно ответить на вопрос, какие сенсорные сигналы приводят к тем или иным движениям во время маневра. Но проведенный труд однозначно предоставляет почву для будущих исследований.
Наблюдения за насекомыми и птицами всегда служили вдохновением для создания человеком летательных аппаратов. Изучение синих мух не будет исключением. Ученые считают, что их наблюдения указывают на то, что для успешной реализации роботизированного или пилотируемого аппарата, способного приземляться на инвертированных поверхностях, необходима плотная интеграция вычислительных и механических процессов. Например, это может быть комбинация схем быстрого распознания изображений и система контроля положения аппарата плюс система стабилизации, которую мухи реализовали посредством колебания тела на лапках.
Когда-то человек мог только мечтать о полетах, сейчас же это реальность. Но мы лишь гости в этой чуждой для нас среде, которая является родной для многих живых организмов, и нам предстоит еще многое понять, открыть и исследовать, прежде, чем мы станем их полноценными соседями в облаках.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята! :)
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, 30% скидка для пользователей Хабра на уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2650 v4 (6 Cores) 10GB DDR4 240GB SSD 1Gbps от $20 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Комментарии (7)

trapwalker
30.10.2019 13:40Интересно натренировать так коптеры на провода аккуратно и надёжно садиться. Можно было бы и заряжаться через токовую петлю на высоковольтных линиях.

OneOfUs
31.10.2019 11:32Представляю новости через несколько лет:
«Крупнейшая авария в электросети оставила без электричества N-дцать населённых пунктов в Энском районе. В аварии виноват рой дронов, севший на провода высоковольтной линии электропередач для дозаправки. Под их тяжестью, провод, не расчитанный на такую нагрузку, оборвался, тем самым лишив электричества N-дцать населённых пунктов...»trapwalker
31.10.2019 11:44Дожить бы до таких новостей=) Забавное будущее.
Мне кажется у энергетиков мгновенно появятся свои такие дроны, которые будут приглядывать за этими птичками, следить чтобы с них ничего не свисало, чтобы на пролёт больше десятка не усаживалось, чтобы не гадили на изоляторы...
NikitOS9
v1000
ну так простая полупетля, но с использованием лапок
NikitOS9
>но с использованием лапок
опора т.е.