

Эволюция подарила нам множество необычных животных. Некоторые из них настолько необычны, что их первооткрывателей называли мошенниками (как это было с утконосами в 1797 году). Но не только фауна может похвастаться нестандартными видами, среди представителей флоры также есть белые вороны, а именно растения, не желающие сидеть на солнечной диете, а предпочитающие сочных насекомых и другую мелкую живность в качестве блюда дня. Ранее мы с вами уже знакомились с исследованием, в котором ученые рассматривали венерину мухоловку и ее маргинальные шипы. Сегодня же нам предстоит узнать, насколько эти шипы чувствительны. Ученые из Цюрихского университета (Швейцария) провели ряд практических опытов, целью которых было измерение скорости реакции венериной мухоловки на прикосновения той или иной силы. Как именно ученые «щекотали» мухоловку, насколько быстро она реагировала, и как полученные результаты поясняют гастрономические предпочтения растения-хищника? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Дабы не растягивать сие повествование, описание венериной мухоловки (которое уже было использовано в предшествующей статье) спрятано под спойлером:
Что такое мухоловка?
Всем нам знакомо понятие фотосинтез, которое обозначает химический процесс преобразования энергии света в энергию химических соединений. Однако далеко не всем растениям по душе солнечная диета, точнее не все могут прокормить себя исключительно этим способом ввиду определенных климатических условий и места произрастания. Хочешь жить — умей вертеться, в данном случае эволюционируй в ночной кошмар всех мух. Другими словами, хищные растения в дополнение к «классическому» питанию получают необходимые вещества (например, азот) еще и переваривая насекомых (или других существ, коим сильно не повезло). Всего насчитывается порядка 630 видов хищных растений, которые произрастают практически во всех регионах планеты и отличаются как по методу охоты, так и по гастрономическим предпочтениям.
По типу ловли добычи хищных растений можно разделить на два основных типа: активные и пассивные. Первые обладают движущимися частями (как наша сегодняшняя героиня мухоловка), а вторые — липкими выделениями на поверхности листьев или ёмкостями, куда жертва сваливается сама.

Библис гигантский
Самыми крупными представителями хищных растений являются библис гигантский и непентес. Библис гигантский напоминает обычный кустарник с очень красивыми цветами, но это растение очень коварно — все его листья и стебли покрыты маленькими волосками (примерно 300 000 штук на 1 листе) и железами, которые выделяют сок. Этот нектар привлекает жертву, которая позарившись на халявную еду сама ею становится, намертво прилипая к волоскам. При этом выделяемый растением сок не просто приманка, но и инструмент переваривания. Жертвами библиса могут быть как насекомые и улитки, так и мелкие амфибии или очень неудачно приземлившиеся птицы.
Симбиотические отношения непентеса и паука, использующего резервуар растения как место для «рыбалки».
Второе название непентеса это кувшиночник, что прямо говорит о методе ловли добычи. У непентеса два вида листьев: обычные и необычные, которые формируют висящий в воздухе кувшин, наполненный нектаром. Края этого кувшина также покрыты нектаром, потому привлеченное его запахом насекомое садится и скатывается внутрь, где и переваривается растением. Но непентес отличается от других растений хищников, он не только серийный убийца, но и хороший друг, если вы тупайя. Этот маленький зверек питается нектаром растения и использует его ловушку-кувшин, как ночной горшок. Продукты жизнедеятельности перевариваются и дают растению необходимые питательные вещества.

Даже в цветении мухоловки присутствует логика: цветы распускаются на длинном стебле подальше от ловушек, дабы уберечь насекомых-опылителей от несчастного случая на производстве.
И, наконец-то, активные хищники. Самым знаменитым среди них является мухоловка. Ее листья на конце тонкого стебля напоминают капкан или раскрытую, голодную и очень зубастую пасть. Внутри капкан покрыт тонкими и очень чувствительными волосками, задев которые жертва активирует захлопывание ловушки. Но такой метод активации не самый эффективный, подумаете вы. А если подул ветер или начался дождь, и волосок был задет случайно? Но не переживайте, ибо механизм этот мухоловка совершенствовала долгие годы эволюции. Чтобы створки сомкнулись необходимо задеть минимум 2 волоска с интервалом не более 20 секунд. При этом процесс переваривания, который протекает внутри капкана, начнется только после повторной стимуляции чувствительных волосков. Таким образом, мухоловка не начнет процесс переваривания, например, упавшего листика, тем самым сохранив энергию для реальной добычи.
Механизм захлопывания ловушки у мухоловок построен на химических и клеточных процессах. Самую важную роль играет тургор тканей, то есть напряженное состояние оболочек живых клеток. Когда жертва стимулирует чувствительные волоски, генерируется потенциал действия за счет ионов калия, который распространяется по долям листа и стимулирует клетки. Когда ловушка открыта, ее доли изогнуты наружу (так жертве будет проще попасть в ловушку), а когда ловушка закрывается, доли изгибаются в обратном направлении. Таким образом формируется внутреннее пространство. Живая жертва не будет стоять на месте, а будет постоянно двигаться, пытаясь выбраться. Но тут как в болоте — чем больше двигаешься, тем быстрее утонешь. Постоянные движения жертвы стимулируют волоски, что приводит к выделению пищеварительного фермента. Если же добыча оказалась умнее и быстрее и смогла сбежать, то закрывшаяся впустую ловушка откроется только через 2-3 дня.
Немаловажный аспект работы ловушек заключается в их недолговечности и деформации. Захлопывание долей листа в холостую приводит к тому, что в последующие разы ловушка будет закрываться медленнее, а угол открытого состояния будет значительно меньше. Как мы с вами понимаем, такие изменения могут сильно отразиться на поимке добычи, а значит и привести к гибели растения ввиду голода. Кроме этого, после нескольких закрытий ловушки достаточно быстро отмирают.
True Facts: Carnivorous Plants (Правдивые факты: хищные растения). Ролик содержит много черного юмора и немного нецензурной лексики.
Важную роль в поимке добычи для мухоловок играют краевые шипы, которые предотвращают побег жертвы из ловушки-желудка. Однако, как и с чувствительными волосками, так и с этими шипами не все так просто.
Исследование, о котором пойдет речь, основано на гипотезах Чарльза Дарвина, который первый описал механизм охоты венериной мухоловки, назвав ее ловушку «ужасной тюрьмой». Он предположил, что шипы важны не только для удержания добычи, но и как своеобразный метод ее «фильтрации». Другими словами, слишком маленькая добыча, питательной пользы от которой будет недостаточно, сможет выбраться между шипами, а слишком крупная добыча просто раскроет доли листа и также сбежит. И в первом, и во втором случае затраты энергии мухоловки на такие виды добычи были бы несопоставимы с полученной в результате энергией от их переваривания. Соответственно, шипы мухоловки позволяют ей ловить добычу определенного размера.
По типу ловли добычи хищных растений можно разделить на два основных типа: активные и пассивные. Первые обладают движущимися частями (как наша сегодняшняя героиня мухоловка), а вторые — липкими выделениями на поверхности листьев или ёмкостями, куда жертва сваливается сама.
Библис гигантский
Самыми крупными представителями хищных растений являются библис гигантский и непентес. Библис гигантский напоминает обычный кустарник с очень красивыми цветами, но это растение очень коварно — все его листья и стебли покрыты маленькими волосками (примерно 300 000 штук на 1 листе) и железами, которые выделяют сок. Этот нектар привлекает жертву, которая позарившись на халявную еду сама ею становится, намертво прилипая к волоскам. При этом выделяемый растением сок не просто приманка, но и инструмент переваривания. Жертвами библиса могут быть как насекомые и улитки, так и мелкие амфибии или очень неудачно приземлившиеся птицы.
Симбиотические отношения непентеса и паука, использующего резервуар растения как место для «рыбалки».
Второе название непентеса это кувшиночник, что прямо говорит о методе ловли добычи. У непентеса два вида листьев: обычные и необычные, которые формируют висящий в воздухе кувшин, наполненный нектаром. Края этого кувшина также покрыты нектаром, потому привлеченное его запахом насекомое садится и скатывается внутрь, где и переваривается растением. Но непентес отличается от других растений хищников, он не только серийный убийца, но и хороший друг, если вы тупайя. Этот маленький зверек питается нектаром растения и использует его ловушку-кувшин, как ночной горшок. Продукты жизнедеятельности перевариваются и дают растению необходимые питательные вещества.
Даже в цветении мухоловки присутствует логика: цветы распускаются на длинном стебле подальше от ловушек, дабы уберечь насекомых-опылителей от несчастного случая на производстве.
И, наконец-то, активные хищники. Самым знаменитым среди них является мухоловка. Ее листья на конце тонкого стебля напоминают капкан или раскрытую, голодную и очень зубастую пасть. Внутри капкан покрыт тонкими и очень чувствительными волосками, задев которые жертва активирует захлопывание ловушки. Но такой метод активации не самый эффективный, подумаете вы. А если подул ветер или начался дождь, и волосок был задет случайно? Но не переживайте, ибо механизм этот мухоловка совершенствовала долгие годы эволюции. Чтобы створки сомкнулись необходимо задеть минимум 2 волоска с интервалом не более 20 секунд. При этом процесс переваривания, который протекает внутри капкана, начнется только после повторной стимуляции чувствительных волосков. Таким образом, мухоловка не начнет процесс переваривания, например, упавшего листика, тем самым сохранив энергию для реальной добычи.
Механизм захлопывания ловушки у мухоловок построен на химических и клеточных процессах. Самую важную роль играет тургор тканей, то есть напряженное состояние оболочек живых клеток. Когда жертва стимулирует чувствительные волоски, генерируется потенциал действия за счет ионов калия, который распространяется по долям листа и стимулирует клетки. Когда ловушка открыта, ее доли изогнуты наружу (так жертве будет проще попасть в ловушку), а когда ловушка закрывается, доли изгибаются в обратном направлении. Таким образом формируется внутреннее пространство. Живая жертва не будет стоять на месте, а будет постоянно двигаться, пытаясь выбраться. Но тут как в болоте — чем больше двигаешься, тем быстрее утонешь. Постоянные движения жертвы стимулируют волоски, что приводит к выделению пищеварительного фермента. Если же добыча оказалась умнее и быстрее и смогла сбежать, то закрывшаяся впустую ловушка откроется только через 2-3 дня.
Немаловажный аспект работы ловушек заключается в их недолговечности и деформации. Захлопывание долей листа в холостую приводит к тому, что в последующие разы ловушка будет закрываться медленнее, а угол открытого состояния будет значительно меньше. Как мы с вами понимаем, такие изменения могут сильно отразиться на поимке добычи, а значит и привести к гибели растения ввиду голода. Кроме этого, после нескольких закрытий ловушки достаточно быстро отмирают.
True Facts: Carnivorous Plants (Правдивые факты: хищные растения). Ролик содержит много черного юмора и немного нецензурной лексики.
Важную роль в поимке добычи для мухоловок играют краевые шипы, которые предотвращают побег жертвы из ловушки-желудка. Однако, как и с чувствительными волосками, так и с этими шипами не все так просто.
Исследование, о котором пойдет речь, основано на гипотезах Чарльза Дарвина, который первый описал механизм охоты венериной мухоловки, назвав ее ловушку «ужасной тюрьмой». Он предположил, что шипы важны не только для удержания добычи, но и как своеобразный метод ее «фильтрации». Другими словами, слишком маленькая добыча, питательной пользы от которой будет недостаточно, сможет выбраться между шипами, а слишком крупная добыча просто раскроет доли листа и также сбежит. И в первом, и во втором случае затраты энергии мухоловки на такие виды добычи были бы несопоставимы с полученной в результате энергией от их переваривания. Соответственно, шипы мухоловки позволяют ей ловить добычу определенного размера.
Основная идея механизма срабатывания ловушки венериной мухоловки заключается в том, что будущей добыче необходимо произвести два касания к чувствительным волоскам растения в течение 30 секунд. Каждое их касание производит потенциал действия*, необходимый для закрытия ловушки. Ученые считали так на протяжении 200 лет, но соответствует ли теория практике?
За годы исследований ученые вывели сценарий событий, который должен (по идее) разворачиваться во время охоты мухоловки:
- голодное растение привлекает насекомых за счет секреции летучих соединений;
- не шибко смекалистое насекомое исследует растение (надеясь на халявную пищу), во время чего касается одного из шести чувствительных волосков, тем самым вызывая потенциал действия (AP от action potential);
- если насекомое в течение 30 секунд не сообразило, что «It’s a trap!», и совершило второе касание, то ловушка захлопывается;
- заключенная между створками ловушки добыча начинает сопротивляться (тщетно, естественно), что приводит к активации синтеза жасмоновой кислоты (C12H18O3) и еще более плотному закрытию створок.
В результате ранее открытые «лепестки» мухоловки превращаются в своеобразный желудок, где и будет перевариваться неудачливая добыча.
В данном труде ученые решили детально рассмотреть процесс преобразования механической стимуляции чувствительных волосков в электрический сигнал. Мы точно знаем, что касание к чувствительным волоскам открывает механочувствительные ионные каналы, но идентифицировать эти каналы пока еще не удавалось.
Пока эти предполагаемые каналы открыты, рецепторный потенциал* (RP от receptor potential) возрастает, и, если отклонение волоска достаточно велико, RP достигает порога, выше которого проявляется AP.
Потенциал действия* — волна возбуждения, котороя проходит по мембране живой клетки в виде кратковременного изменения мембранного потенциала.
Рецепторный потенциал* — возникает при деполяризации поверхностной мембраны рецептора, ввиду воздействия на него раздражителя.
Потенциал покоя* — мембранный потенциал возбудимой клетки в невозбужденном состоянии.
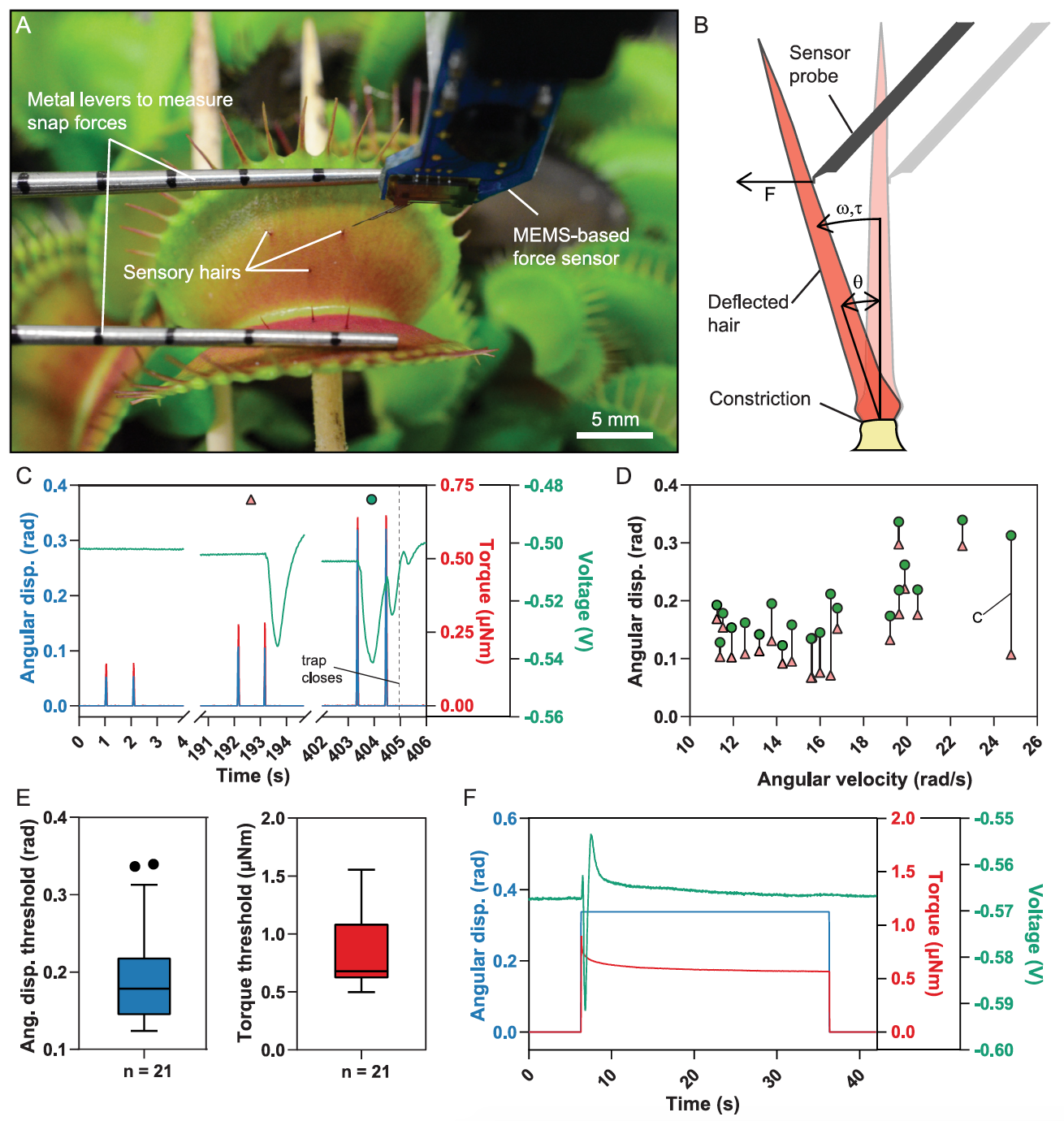
Изображение №1
Ранее уже были попытки связать механические стимулы и генерацию потенциала действия, но не было подходящих инструментов для точных измерений. В случае рассматриваемого нами сегодня исследования, ученые применили датчик силы на основе MEMS*, установленный на микророботной системе, для точного контроля скорости и амплитуды отклонения, а также одновременного измерения приложенной силы (1A и 1B).
MEMS* (микроэлектромеханические системы) — устройства, объединяющие в себе микроэлектронные и микромеханические компоненты.
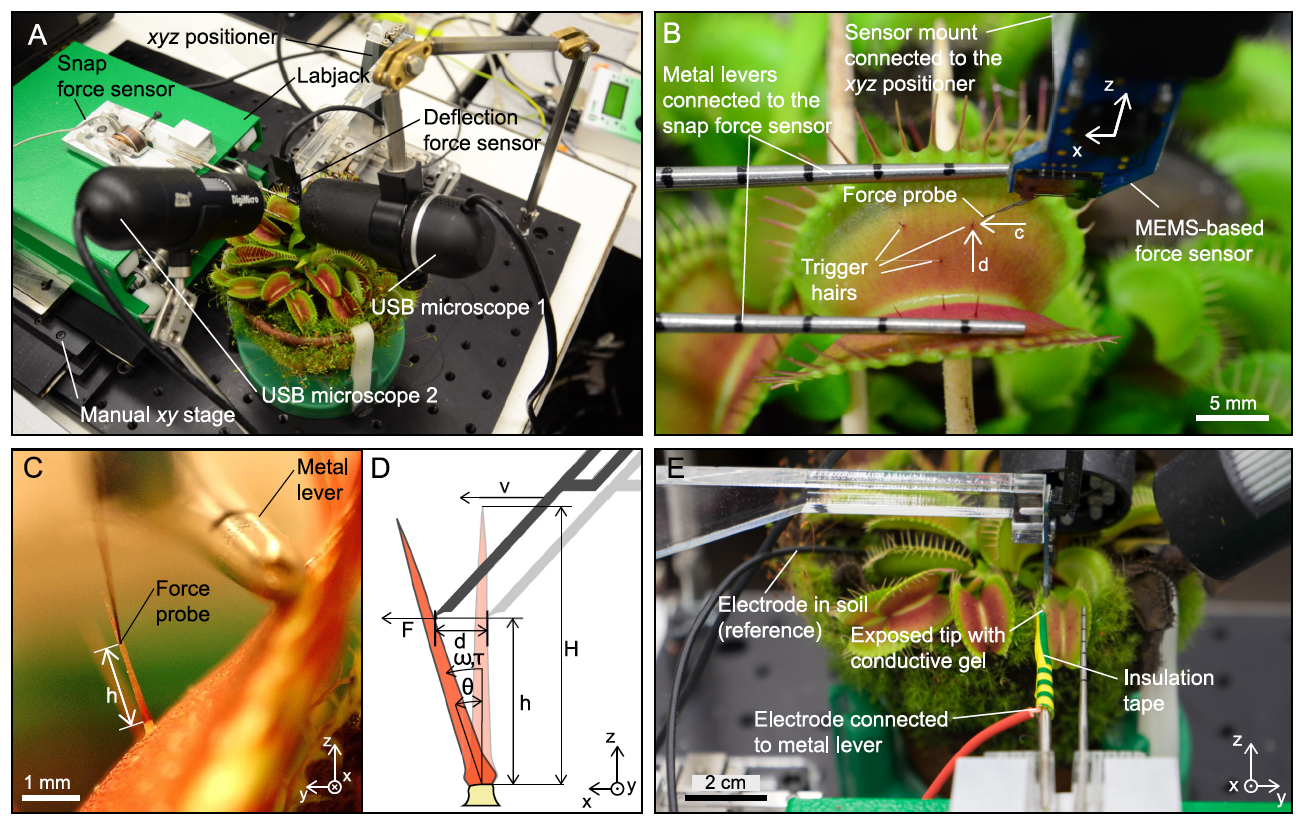
Изображение №2
Таким образом, ученые смогли точно определить диапазон параметров, в котором отклонение волосков приводит к закрытию ловушки, в то время как второй датчик силы измерял сгенерированное мгновенное усилие (изображение выше).
Результаты исследования
В процессе опытов ученые учли, что пауки, мухи и муравьи (обычная добыча для мухоловки) будут вызывать достаточно быстрое отклонение чувствительных волосков. В связи с этим микророботная система была задействована на полной скорости, чтобы имитировать эти раздражители. Это привело к высокой начальной угловой скорости в диапазоне от 10 до 20 рад/с.
В ранее проведенных исследованиях было установлено, что муравьи отклоняют чувствительные волосы с угловой скоростью 0.25 – 7.8 рад/с, но медленнее мух. Следовательно, используемые параметры выше реальных, что хорошо подходит для оценки возможностей венериной мухоловки.
При таких высоких скоростях продолжительность прогиба значительно короче, чем у других вовлеченных зависящих от времени факторов, таких как затухание RP и расслабление чувствительных волосков (1F).
Учет углового, а не линейного отклонения позволил скорректировать различия в высоте контакта сенсорного зонда относительно места сужения сенсорного волоска, а также различия, касающиеся разных геометрий сенсорного волоска (1B, 2C и 2D). Следовательно, одно отклонение может быть аппроксимировано дискретным увеличением углового смещения, и запуск AP в основном зависит от величины углового смещения.
Одно отклонение было определено как комбинация углового смещения «вперед-назад», подобное тому, что происходит при касании к волоску добычей. Каждое измерение состояло из двух последующих прогибов с промежутком в 1 с между ними до заданного углового смещения ?. Если ловушка не закрылась, следовал этап ожидания (2 минуты), чтобы убедиться, что RP полностью сброшен. Ожидание в течение именно двух минут, а не больше/меньше, обусловлено тем, что серия более ранних экспериментов продемонстрировала, что при температуре ниже 30 °C для быстрого и полного закрытия ловушки необходимы два прогиба волоска в течение 30–40 с.
После этапа ожидания процедуру повторяли с увеличением угловых смещений до тех пор, пока не срабатывал механизм закрытия ловушки (1С и 1D). Это происходило, когда был достигнут средний порог смещения ? = 0.18 рад или средний порог крутящего момента ? = 0.8 мкН·м (n = 21).
Ученые отмечают, что ни разу не было зафиксировано закрытие ловушки, если ? был ниже 0.12 рад и ? был ниже 0.50 мкН·м (1Е). Следовательно, эти показатели являются нижним пределом углового отклонения, который необходим для запуска закрытия ловушки в условиях данного эксперимента. Также удалось определить, что для срабатывания ловушки насекомое должно прикладывать силу (F) в 0.5 мН вблизи кончиков волосков или 5 мН вблизи основания.
Измерения AP обеспечили связь между отклонением чувствительных волосков и закрытием ловушки. Когда два последовательных отклонения были значительно ниже порога смещения (? << 0.12 рад), AP не наблюдался. Для амплитуд отклонения вблизи порога смещения (? < 0.12 рад) после второго отклонения был выявлен однократный AP.
Это указывало на то, что оба отклонения волосков вносят вклад в RP и что порог индукции AP был достигнут только при втором отклонении. Как и ожидалось, однократного AP было недостаточно для срабатывания ловушки.
Предположение, что каждое прикосновение вызывает AP, реально только в том случае, если отклонение сенсорных волосков превышает порог смещения. Если это так, то два AP (по одному на каждое отклонение волоска) генерировались и приводили к закрытию ловушки (1С).
Двойное отклонение чувствительных волосков, приводящее к двум AP и закрытию ловушки.
Эти результаты предполагают, что быстрое отклонение чувствительных волосков увеличивает RP до определенного уровня, который зависит от амплитуды углового отклонения. RP могут складываться и могут вызывать AP после нескольких отклонений, если они ниже порога отклонения. Тем не менее, генерация одного AP за касание происходит только в том случае, если отклонение чувствительных волосков выше порога отклонения.
Другими словами, потенциал действия будет генерироваться только тогда, когда чувствительный волосок был «потревожен» достаточно сильно. Но однократного потенциала действия недостаточно, чтобы спровоцировать закрытие ловушки. Это вполне вписывается в рамки теории о том, что мухоловки не будут тратить силы на закрытие ловушки при каждом касании чего-то или кого-то к их волоскам. В противном случае может быть много холостых срабатываний ловушки без какой-либо энергетической (т.е. питательной) отдачи (добыча успела сбежать, добыча была слишком мала или это вовсе была не добыча, а мусор).
Учитывая, что увеличение RP при многократных отклонениях является аддитивным, логично будет предположить, что устойчивое (длительное) смещение волоска может иметь аналогичный эффект.
Для проверки этой гипотезы ученые отклонили чувствительный волосок мухоловки за пределы порога углового смещения и зафиксировали его в таком положении на 30 секунд (1F). Однако это не привело к срабатыванию ловушек.
Изначальное смещение вызвало однократный AP, после чего напряжение быстро вернулось к базовому значению, несмотря на то, что волоски оставались отклоненными.
Если бы устойчивое смещение вносило свой вклад в RP, оно должно было бы остаться выше порогового значения, и в этом случае ожидалась серия AP. Дополнительные анализы показали, что угловое смещение действительно играет важную роль в RP, но статическое отклонение волосков не вносит никакого вклада в этот процесс.
Любопытно, что в ранее проводимых опытах ситуация обстояла иначе: однократное смещение волосков приводило к закрытию ловушки. Однако, это было не целостное однократное смещение, а множество мелких смещений, так как колебания, которые сопровождаются ручным отклонением волоска, вероятно, больше, чем порог углового смещения.
Следующий этап исследования заключался в создании модели накопления электромеханического заряда (ECB от electromechanical charge buildup), которая на удивление предсказывает закрытие ловушек от однократного касания.
Взяв за основу полученные в ходе опытов данные, ученые разработали простую модель для изучения пределов углового смещения и скорости, в рамках которых ловушки будут реагировать.
В ECB модели механическое отклонение приводит к накоплению заряда RP как функции угловой скорости ? и смещения ?, в то время как заряды непрерывно рассеиваются. Если накопленные заряды превышают определенное пороговое значение QthRP, то возникает AP. Дополнительно был имплементирован рефрактерный период tRP, представляющий временной интервал, необходимый для того, чтобы мог быть активирован еще один AP.
В результате модель предсказала, что если отклонения слишком быстрые и/или слишком малые, то может потребовать более двух таких отклонений, чтобы вызвать закрытие ловушки (красная область на 3А).

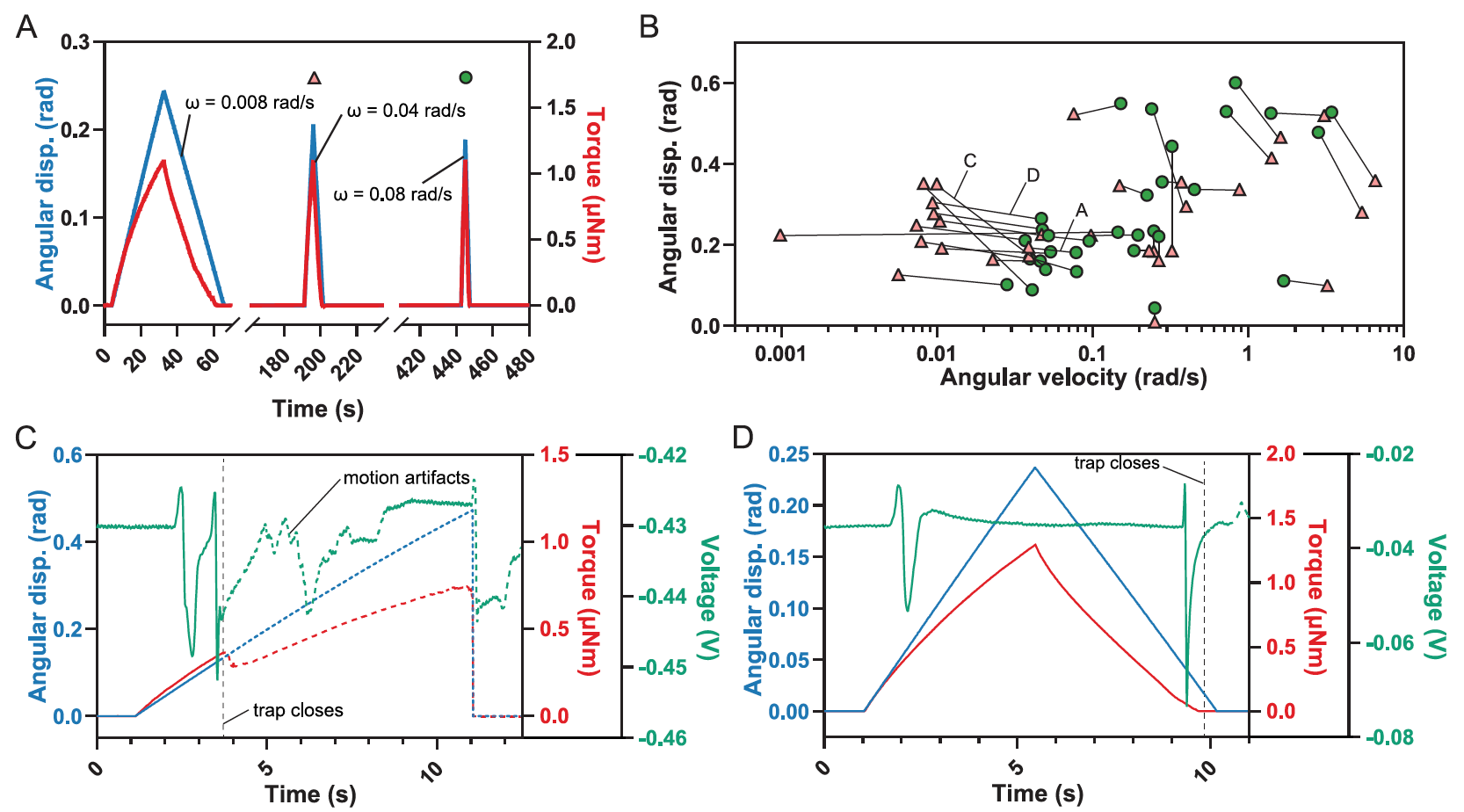
Изображение №3
Это объясняется тем, что одного отклонения недостаточно для активации AP (3В), что и показали практические опыты (1С, в центре).
Аналогично, модель показала, что очень низкие угловые скорости (? < 0.04 рад/с) не могут заполнить RP.
Но самым неожиданным результатом моделирования стало предсказание диапазона промежуточных угловых скоростей (0.04 рад/с < ? < 10 рад/с), при которых одного отклонения достаточно, чтобы активировать два или более AP, которые необходимы для закрытия ловушки (3B).
Однако такое предсказание противоречит представлению о том, как работает механизм ловушки венериной мухоловки. Потому ученые решили проверить, можно ли реализовать эту модельную ситуацию на практике.
Однократное отклонение чувствительных волосков, приводящее к закрытию ловушки.
Удивительно, но закрытие ловушки действительно происходило при одиночном отклонении волосков, если угловые скорости были достаточно низки.
Чтобы сузить диапазон значений, в котором это происходит, ученые неоднократно отклоняли одни и те же сенсорные волоски с различными угловыми скоростями, пока ловушка не закрылась. Между двумя последовательными отклонениями был промежуток в 2 минуты, пока ловушка не восстановится и не исчерпается RP.
Нижняя граница угловой скорости ?, необходимой для закрытия ловушки при одном отклонении, определялась путем постепенного увеличения угловой скорости после каждого отклонения (n = 17). Начальная скорость была ниже 0.009 рад/с, так как при такой скорости никогда не наблюдалось закрытия ловушки.
Изображение №4
Последующие одиночные отклонения волосков выполнялись при постепенном увеличении скорости, пока ловушка не срабатывала (4А).
Аналогичным образом определяли верхнюю границу, начиная со скорости ? > 3 рад/с с последующим пошаговым снижением (n = 9).
Кроме того, была выполнена еще одна серия экспериментов с одним отклонением (n = 5), в которой скорость силового зонда поддерживалась постоянной, что приводило к промежуточной угловой скорости между 0.2 и 0.4 рад/с, в то время как угловое смещение ? постепенно увеличивалось во время последующих отклонений, чтобы получить нижнюю границу ?, необходимую для срабатывания ловушки в условиях однократного отклонения волосков.
Все однократные отклонения, приводящие к закрытию ловушки, вместе с предшествующими стимулами, для которых закрытие ловушки не происходило, определяли область, где одиночное отклонение вызывает закрытие (4B).
Однократное отклонение может вызвать закрытие ловушки при промежуточных угловых скоростях отклонения (0.03 рад/с ? ? ? 4 рад/с). Но одного отклонения при более низких или высоких скоростях будет недостаточно.
Эксперименты, где скорость отклонения волосков была достаточно низкой, показали, что срабатывание ловушки происходит как при приближении датчика к основанию волоска, так и при удалении датчика от него.
Однократное отклонение чувствительных волосков, приводящее к двум AP во время начального изгиба, приводя к закрытию ловушки.
Когда ловушка закрывалась во время приближения зонда, во время отклонения волосков наблюдалось два последовательных потенциала действия (4C). Когда ловушка закрывалась во время удаления зонда, один АР возникал в момент отклонения волоска, а второй уже после удаления зонда, когда волосок возвращался в исходное положение (4D). В обоих случаях второй АР приводил к немедленному закрытию ловушки.
В заключение ученые решили измерить силу закрытия ловушки (5А).
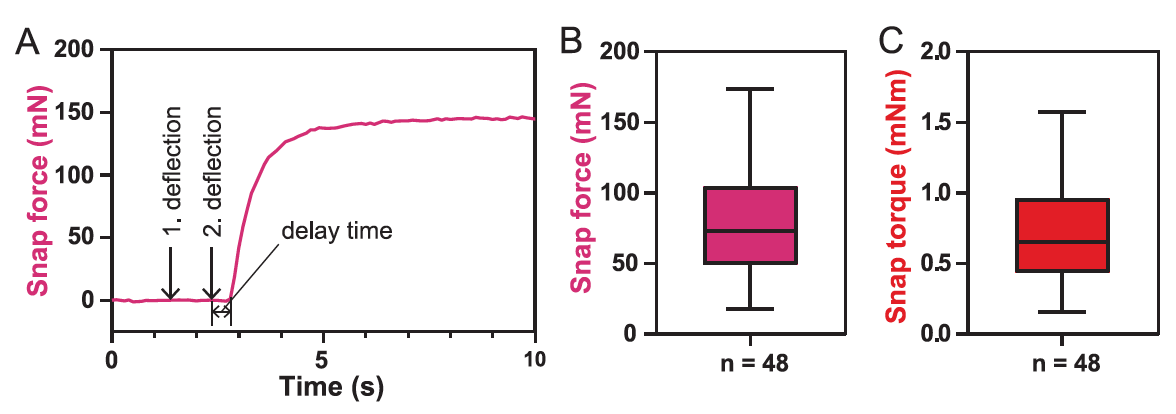
Изображение №5
Для 48 ловушке среднее значение силы захвата Fclose составило 73 мН (5В), а диапазон значений был от 18 до 174 мН.
Данные значения ниже тех, что были опубликованы ранее (140-150 мН). Тому есть объяснение: в данном случае измерялась сила в начале закрытия ловушки, а в предыдущих наблюдениях измерялась сила, воздействующая на края створок листа-ловушки по завершению закрытия.
Поскольку измеренная сила сильно зависит от положения датчика, а также от ориентации и размера листа, крутящий момент закрытия вокруг середины листа ?close со средним значением 0.65 мН·м является лучшей величиной для описания силы закрытия ловушки (5C). Время задержки, то есть время между механическим стимулом и началом закрытия ловушки, составило 0.6 ± 0.3 с.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
Венерина мухоловка не бегает, как гепард, не обладает смертоносным ядом, как кобра, и уж точно не слышит свою добычу, как сова. Но это растение действительно является одним из лучших хищников на планете, так как его механизм охоты работает, как часы.
В данном труде ученые смогли определить какими должны быть те или иные параметры, чтобы ловушка сработала. Как оказалось, в большинстве случаев для срабатывания капкана жертва должна совершить две последовательные ошибки: первая — дотронуться до сенсорного волоска мухоловки; вторая — сделать это еще раз.
Повторное отклонение волоска от его изначального положения приводит к генерации второго потенциала действия, который запускает процесс закрытия ловушки. Тем не менее, моделирование показало, что при определенных условиях может быть достаточно и одного касания, но в реальности это скорее всего маловероятно.
Другими словами, может ли венерина мухоловка послужить в качестве охотника на комаров? Нет, ибо эти кровососы очень маленькие и слишком шустрые. Но вот мух или улиток мухоловка не жалеет.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?





zloddey
Извините
anonymous
Мне статья понравилась.
zloddey
Мне тоже, но уж больно напрашивалась аналогия...