

Когда нам что-то не нравится, мы всячески пытаемся избегать какого-либо контакта с этим. Если же обычная неприязнь сменяется страхом, то желания контактировать с его источником еще меньше. Порой нам даже смотреть не хочется на то, что нас пугает. Все процессы, протекающие в организме человека, в той или иной степени контролируются мозгом. Одернуть руку от горячей поверхности, повернуть голову в сторону неожиданного громкого звука, определить испорченность продукта по его запаху — все это результат работы мозга, направленной на сохранение жизни и здоровья всего организма. Многие из механизмов работы ЦНС вполне неплохо изучены, но как именно мозг подталкивает нас отвести взгляд от чего-то страшного пока неясно. Ученые из Токийского университета (Япония) провели исследование, в котором им удалось раскрыть это тайну, а помогли им в этом плодовые мушки. Почему мы отводим взгляд от пугающих нас вещей, как этим процессом управляет мозг, и где могут пригодиться результаты данного исследования? Ответы на эти вопросы мы найдем в докладе ученых.
Основа исследования
Окружающая среда полна опасностей, а потому адаптация к ее непредсказуемым условиям является одним из краеугольных камней выживания любого живого организма. Погодные условия, хищники, потенциальные партнеры, источники пищи — все эти аспекты жизни животного должны каким-то образом восприниматься и дифференцироваться. Тут на помощь приходят сенсорные системы — слух, зрение, обоняние и т.д.
Животные, находящиеся под угрозой, не только демонстрируют реакции бегства и сердечные реакции, но также резко модулируют широкий спектр сенсорных реакций, включая ноцицепцию, слух и зрение. Из них зрение представляет собой самый богатый источник информации для дневных животных об окружающей среде. Следовательно, его модуляция в опасной ситуации, вероятно, играет жизненно важную роль.
Исследования животных выявили нейронные механизмы, с помощью которых угроза вызывает зрительное отвращение. У рыб, например, звуковой сигнал, который сам по себе вызывает реакцию испуга, также усиливает реакцию испуга на визуальный сигнал. Эта регуляция может быть достигнута за счет возбуждающей конвергенции механических и зрительных сигналов ниже по течению от тектума зрительного нерва. В дополнение к конвергенции возбуждения известно, что зрительные цепи, такие как первичная зрительная кора, латеральное коленчатое тело, верхнее двухолмие и даже клетки сетчатки или их эквиваленты, находятся под сильным влиянием нейромодуляции. Однако то, как угроза может регулировать зрительные реакции, в частности, структуру мозга, регулируемую угрозой, и молекулярный нейромодулирующий механизм такой регуляции, остается пока в значительной степени неизвестным.
Для поисков ответов на эти вопросы необходим модельный организм, достаточно хорошо изученный, обладающий схожей зрительной системой с точки зрения анатомии и реактивного поведения. Таким организмом является Drosophila melanogaster, т. е. плодовая мушка (дрозофила).
Зрительная система мошек достаточно сложна, но неплохо изучена. При этом она подвергнута богатому спектру модуляции (влиянию внешних и внутренних факторов). Одним из наиболее известных примеров является эффект локомоции, сохраняющийся у разных видов, но другие включают эффекты запаха пищи и сексуального возбуждения. В то время как известные формы зрительной нейромодуляции основаны на функциональном эквиваленте норадреналина, называемом октопамином, дрозофилы содержат гораздо больше эволюционно законсервированных нейромодуляторов. Кроме того, дрозофила, находящаяся в опасной ситуации, проявляет признаки внутреннего состояния, вызванного угрозой. В частности, мухи, подвергшиеся визуальным угрозам, бегают, прыгают, замирают и подавляют кормление. Точно так же мухи, подвергающиеся воздействию серии дуновений воздуха (далее дуновения), усиливают локомоцию и обонятельную реакцию испуга, при этом локомоция регулируется Dop1R1. Также наблюдается брадикардия в случае реакции замирания и тахикардия в случае реакции бегства.
В рассматриваемом нами сегодня труде ученые провели серию опытов с участием дрозофил, в ходе которых было установлено, что дуновения, которые вызывают у дрозофилы признаки внутреннего состояния, временно вызывают отвращение (отвод взгляда) к нейтральному маленькому визуальному объекту.
Далее ученые идентифицировали нейропептид тахикинин и один кластер экспрессирующих его нейронов («нейроны Tk-GAL42 ∩ Vglut»), которые отвечают за стробирование* зрительного отвращения. Эксперименты с визуализацией кальция показали, что нейроны Tk-GAL42 ∩ Vglut кодируют дуновения как повышенную активность. Неожиданной находкой стало то, что эти нейроны кодируют визуальный объект как усиление θ-колебаний только тогда, когда дуновения даются заранее, что причинно связано с визуальным отвращением.
Сенсорное стробирование* (гейтинг, фильтрация) — нейронные процессы фильтрации избыточных или нерелевантных стимулов из всех возможных стимулов окружающей среды, достигающих мозга. Сенсорная фильтрация предотвращает перегрузку информацией в высших корковых центрах мозга. Может проявляться в различных формах посредством изменений как восприятия, так и ощущения, на которые влияют различные факторы (возбуждение, недавнее воздействие стимула и избирательное внимание).
Результаты исследования
Изображение №1
Для дрозофилы небольшой визуальный объект может сигнализировать о присутствии либо потенциального партнера, либо хищника, и, следовательно, его этологическая ценность зависит от контекста. Соответственно, маленький объект обычно не вызывает никакой реакции или вызывает лишь легкое отвращение. Если же объект сочетается с запахом пищи или сексуальным возбуждением, то реакция будет противоположной. Следовательно, отвращение к маленькому объекту усиливается в случае опасной ситуации.
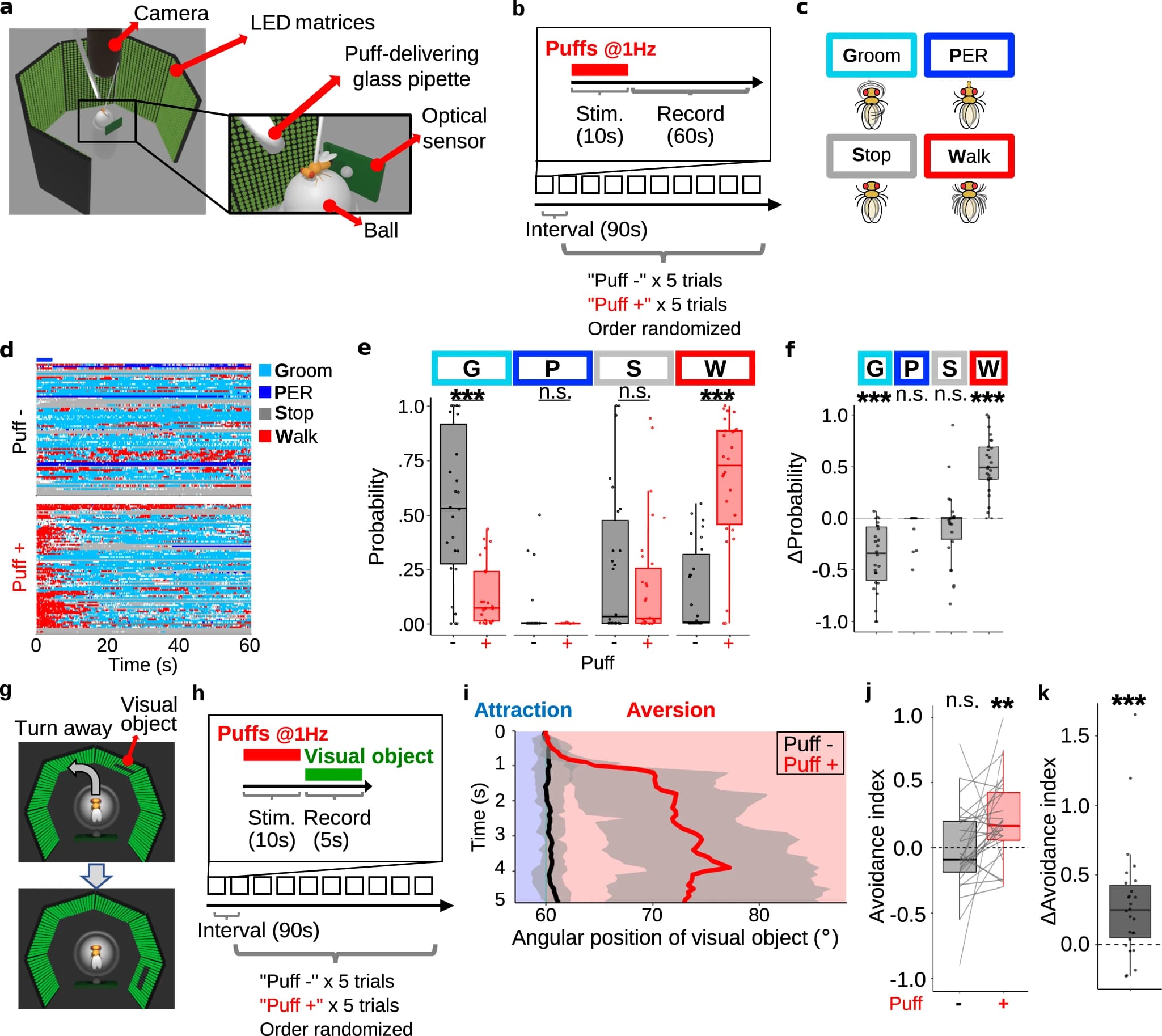
Чтобы проверить эту гипотезу, ученые воспользовались симулятором ходьбы, в котором легко представить дуновения как механические угрозы и визуальный объект в выбранном месте и в выбранное время (1a). Поскольку в предыдущих исследованиях сообщалось об увеличении локомоции как об одном из основных проявлений внутреннего состояния, вызванного угрозой, ученые сначала попытались подтвердить, что дуновения действительно увеличивают локомоцию в рамках проводимого опыта (1b).
Оценка скорость ходьбы мухи по движению мяча (1a) показала, что скорость ходьбы мухи мгновенно увеличивалась при воздействии дуновения (10 раз по 500 мс с частотой 1 Гц). Увеличения числа дуновений приводило к увеличению скорости движения мухи.
Чтобы лучше охарактеризовать поведенческие изменения, вызванные дуновениями, ученые количественно оценили умывание, замирание, рефлекс разгибания хоботка (PER от proboscis extension reflex) и ходьбу с помощью специально написанного программного обеспечения сверточной нейронной сети (1c). Этот анализ показал, что умывание и остановка преобладали перед дуновениями (верхняя панель на 1d). Однако после дуновений это поведение сменилось ходьбой (нижняя панель на 1d), особенно в течение первых 5 секунд (синяя полоса над верхней панелью на 1d). Это наблюдение было подтверждено, когда ученые количественно оценили вероятность каждого поведения в течение первых 5 секунд записи (1e), а также их различия между испытаниями «с дуновениями» и «без дуновений» (1f). Более того, в эксперименте с замкнутым контуром, в котором дуновения применялись только тогда, когда муха двигалась в определенных направлениях, мухи явно избегали движения в этом направлении. Все эти наблюдения подтверждают гипотезу о том, что дуновения воздуха представляют собой механическую угрозу для мух.
Затем было проверено, вызывают ли дуновения воздуха визуальное отвращение, т. е. отведения взгляда. С этой целью перед мухой ставили небольшой визуальный объект, изначально расположенный под углом 60 градусов вправо или влево по отношению к мухе случайным образом, который меняет свое положение в соответствии с моделью ходьбы мухи (замкнутый цикл) (1g). Одно испытание состояло из 90-секундного интервала, 10-секундных дуновений воздуха и 5-секундной записи и повторялось для каждой мухи 10 раз с или без дуновений в случайном порядке (1h).
Видео №1
В случае отсутствия дуновений направленное движение мухи к визуальному объекту или от него не было очевидным (черная линия на 1i; видео №1). Когда же объект демонстрировался сразу после дуновения, мухи явно оборачивались к нему спиной (красная линия на 1i; видео №1).
Этот эффект, однако, может быть вызван просто тем, что повышенная двигательная активность усиливает отвращение, которое слишком слабо, чтобы его можно было обнаружить без предшествующих дуновений. Чтобы решить эту проблему, ученые рассчитали индекс избегания, определяемый линейным расстоянием, пройденным от объекта, деленным на общее линейное пройденное расстояние. Этот показатель увеличился после дуновений (1j). Это свидетельствует о том, что повышенная локомоция не влияет на реакцию отвращения/избегания. Вышеописанные реакции наблюдались для всех исследованных особей (1k). Следовательно, дуновения воздуха служат для дрозофилы механической угрозой, вызывающей отвращение к маленькому визуальному объекту.
Изображение №2
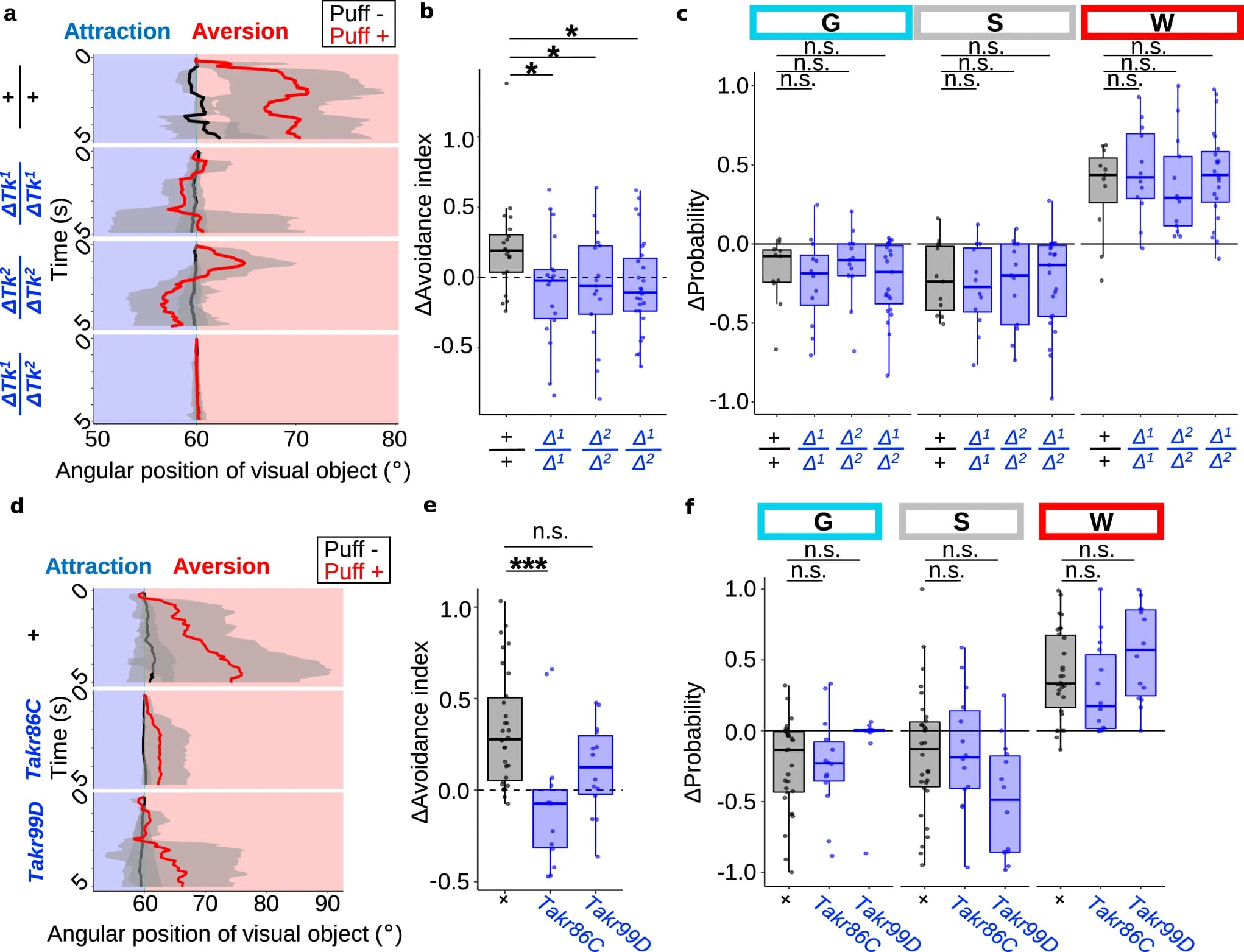
Чтобы исследовать молекулярный механизм, лежащий в основе зрительного отвращения к потокам воздуха, ученые провели скрининг набора нейропептидных нулевых мутантов, сгенерированных CRISPR. Это выявило несколько линий, наиболее заметной из которых была мутантная линия тахикинина (Tk), которая не смогла устранить визуальное отвращение при увеличении локомоции. Чтобы дополнительно подтвердить необходимость Tk, ученые протестировали дополнительные нулевые мутации (Δtk1 и Δtk2).
Интересно, что повышенная локомоция при дуновении воздуха существенно не отличалась между диким типом и этими мутантами (2c). Это указывает на то, что Tk специфически необходим для стробирования зрительного отвращения, но не для повышенной локомоции. Мутанты Tk продемонстрировали оптомоторную реакцию и отвращение к дуновениям в степени, сравнимой с таковой у мух дикого типа. Это позволяет предположить, что эти мутанты сохраняют основные зрительные и двигательные функции.
Чтобы получить дополнительные доказательства того, что Tk необходим для управления зрительным отвращением, ученые протестировали мутанты рецепторов Tk. В геноме мухи идентифицированы Takr86C и Takr99D. Таким образом, была протестирована предполагаемая инсерционная мутация с потерей функции в Takr99D и делеционная мутация с потерей функции в Takr86C. У мутанта Takr86C (2d, 2e), фенокопирующего мутацию Tk (2a, 2b), зрительная аверсия (отвод взгляда) была подавлена. Аналогичная тенденция наблюдалась и для мутанта Takr99D, но не на статистически значимом уровне (2a, 2b). Также в соответствии с фенотипом мутаций Tk (2c), оба мутанта Takr показали повышенную локомоцию (2f). Эти данные подтверждают идею о том, что Tk особенно необходим для стробирования зрительного отвращения, но не для усиления локомоции.

Изображение №3
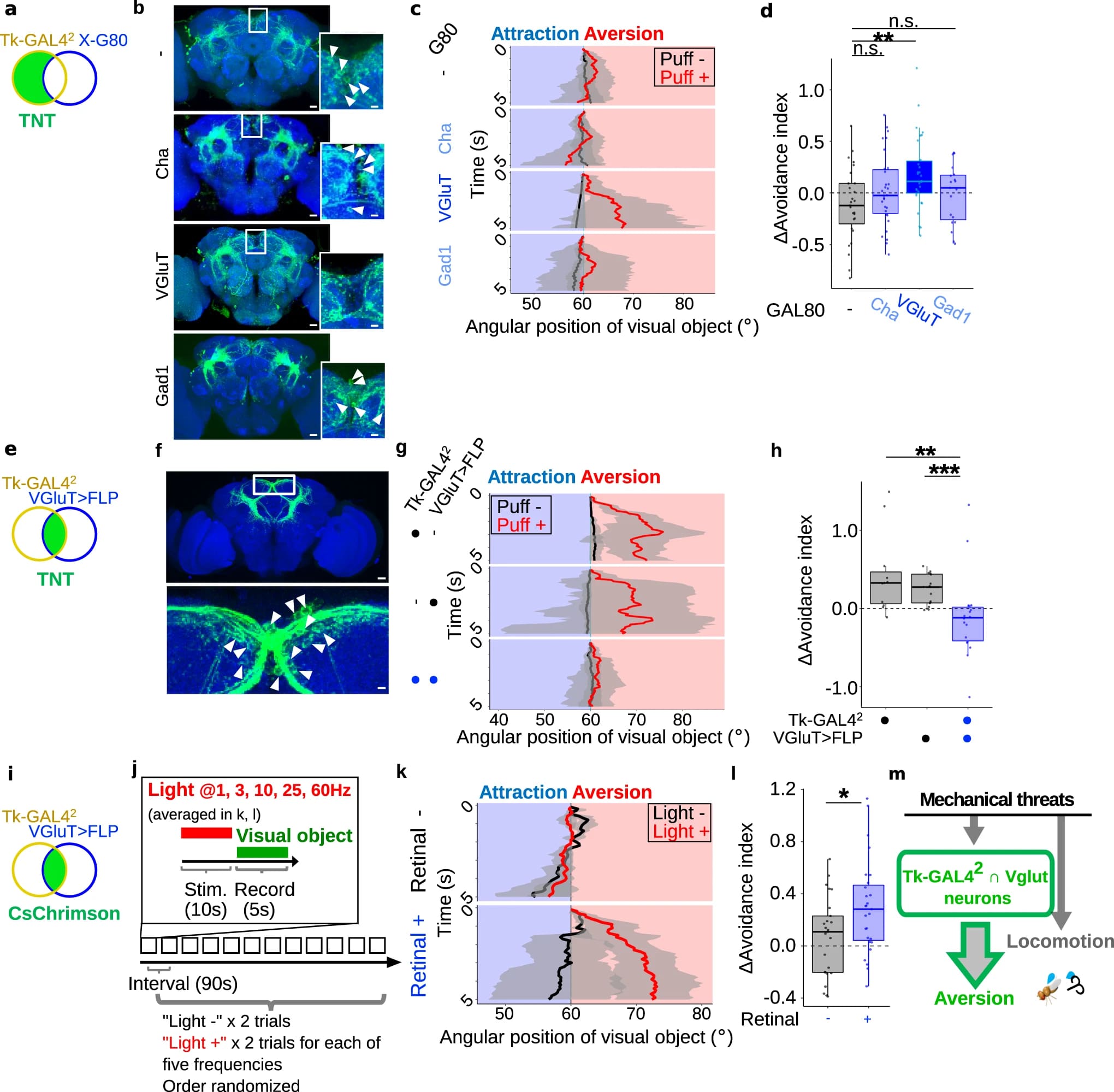
Затем ученые попытались идентифицировать Tk-экспрессирующие нейроны, ответственные за стробирование визуального отвращения. С этой целью они экспрессировали тетанотоксин (TNT от tetanus toxin) с использованием драйверов GAL4 под контролем разных промоторов Tk, каждый из которых приводил к частично перекрывающимся, но разным паттернам экспрессии (3a).
Было обнаружено, что визуальное отвращение к дуновениям подавлялось у мух, у которых TNT экспрессировался одним из этих драйверов (Tk-GAL42), но не другими драйверами. Это позволяет предположить, что эта линия помечает ответственные нейроны (3b, 3c). Экспрессия EGFP и RedStinger с помощью Tk-GAL42 пометила набор из примерно 50 нейронов, разбросанных по всему мозгу. Подавление нейронов Tk-GAL42 не смогло нарушить усиленную локомоцию (3d), предполагая, что нейроны Tk-GAL42 необходимы для визуального отвращения, но не для усиленной локомоции.
Затем необходимо было проверить, может ли искусственная активация данных нейронов заменить воздействие дуновениями воздуха. С этой целью нейроны Tk-GAL42 были активированы путем освещения головы мухи, экспрессирующей светозависимый катионный канал CsChrimson (3e). Одно испытание состояло из 90-секундного интервала, 10-секундной фотоактивации и 5-секундной записи (визуальный объект предъявлялся вскоре после окончания фотоактивации) и повторялось для каждой мухи 6 раз с или без яркого света случайным образом. Действительно, фотоактивация нейронов Tk-GAL42 сама по себе вызывала зрительное отвращение (две верхние панели на 3f).
Это наблюдение было подтверждено, когда была рассчитана разница в индексах избегания между испытаниями с и без воздействия света (два графика слева на 3g). Важно отметить, что этот эффект активации не проявлялся на фоне Tk-KO (нокаут Tk) (нижняя панель на 3f; правый график на 3g). Это позволяет предположить, что фотоактивация нейронов Tk-GAL42 стробирует зрительное отвращение через Tk. В соответствии с данными мутантов Tk и супрессией нейронов Tk-GAL42, фотоактивация нейронов Tk-GAL42 не увеличивала локомоцию на статистически значимом уровне (3h).
Эти данные в совокупности свидетельствуют о том, что нейроны Tk-GAL4 необходимы и достаточны для управления визуальным отвращением, но не для усиления локомоции, и что эти нейроны осуществляют такую функцию через Tk.
Изображение №4
Ученые также пытались сузить подмножество нейронов Tk-GAL42, которые отвечают за стробирование визуального отвращения.
Растущее количество данных свидетельствует о том, что нейропептидергические нейроны обычно совместно экспрессируют один или несколько дополнительных малых нейротрансмиттеров, включая экспрессирующие Tk нейроны в мозге дрозофилы.
Таким образом, ученые пришли к выводу, что объединение Tk-GAL42 с GAL80 (супрессором GAL4), управляемым маркерами малых нейротрансмиттеров (4a), может маркировать различные подмножества нейронов Tk-GAL42. Действительно, эта стратегия привела к маркировке частично перекрывающихся, но различных субпопуляций нейронов Tk-GAL42 (4b).
В частности, GAL80, управляемый маркером глутаматергических нейронов (VGluT), но не маркерами холинергических (ChAT) или ГАМКергических (Gad1) нейронов, подавлял мечение кластера нейронов в верхнем медиальном протоцеребруме (белые стрелки указывают на тела клеток на 4b). Примечательно, что VgluTGAL80 восстанавливал зрительное отвращение к дуновениям, нарушенное TNT, управляемым Tk-GAL42 (4c, 4d). Это позволяет предположить, что VGluT-GAL80 отмечает подмножество нейронов Tk-GAL42, ответственных за стробирование визуального отвращения.
VgluTGAL80 сам по себе не влиял на визуальное отвращение к дуновениям, так как мухи, несущие VGluT-GAL80 в дополнение либо к Tk-GAL42, либо к UAS-TNT, демонстрировали зрительное отвращение, сравнимое с мухами, несущими только Tk-GAL42 или UAS-TNT.
Во всех испытуемых группах наблюдалось увеличение локомоции, сравнимое с таковым в контрольной группе, что подтверждает более ранний вывод (3d, 3h) о том, что нейроны Tk-GAL42 необходимы для повышенной локомоции.
Чтобы более непосредственно исследовать функцию подмножества VGluT+ нейронов Tk-GAL42, ученые затем воспользовались перекрестной стратегией, в которой бинарная система GAL4/UAS была объединена с техникой рекомбинации Flippase (FLP). С помощью этой стратегии ученые стремились ограничить экспрессию TNT клетками, в которых активны как Tk-GAL42, так и VGluT-FLP (зеленая область на 4e).
Иммуногистохимия показала, что пересечение Tk-GAL42 ∩ VgluT действительно маркирует один кластер из 20-30 нейронов в верхнем медиальном протоцеребруме (белые стрелки указывают на тела клеток на 4f), паттерн дополняет паттерн комбинации Tk-GAL42 ∩ VGluT-GAL80 (4b).
Далее эти нейроны были обозначены как нейроны Tk-GAL42 ∩ Vglut. Иммуногистохимия нейронов Tk-GAL42 ∩ Vglut у самок выявила отличную, но частично перекрывающуюся морфологию нейритов по сравнению с самцами.
Чтобы выяснить, необходимы ли нейроны Tk-GAL42 ∩ Vglut для визуального отвращения к дуновениям, TNT экспрессировали в этих нейронах. Эта манипуляция подавляла зрительное отвращение к дуновениям по сравнению с контрольными группами (4g, 4h). Это позволяет предположить, что нейроны Tk-GAL42 ∩ Vglut необходимы для управления зрительным отвращением.
Поскольку искусственная индукция нейронов Tk-GAL42 заменила дуновения воздуха при стробировании визуального отвращения (3e-3g), подобная картина может быть и в случае нейронов Tk-GAL42 ∩ Vglut. После фотоактивации GAL42 ∩ Vglut, экспрессирующих CsChrimson (4i, 4j), зрительное отвращение было стробировано в тестовой группе, но не в контрольной (4k, 4l). Фотоактивация нейронов Tk-GAL42 ∩ Vglut не увеличивала локомоцию на статистически значимом уровне, что указывает на специфическую роль этих нейронов именно в стробировании зрительного отвращения. Следовательно, активация нейронов Tk-GAL42 ∩ Vglut связана с отведением взгляда, но не связана с усилением локомоции (4m).

Изображение №5
На следующем этапе исследования ученые попытались установить, кодируют ли нейроны Tk-GAL42 ∩ Vglut дуновения воздуха и зрительный объект, и если да, то как (5a). С этой целью была выполнена визуализация активности нейронов Tk-GAL42, экспрессирующих GCaMP7f, у зафиксированной мухи при воздействии на нее дуновений воздуха и визуального объекта (5b).
Чтобы зарегистрировать ответ нейронов на только визуальный объект, только дуновения или на дуновения, сопровождаемые визуальным объектом, были протестированы эти три условия с интервалами для одной и той же мухи (5c). Эксперимент показал, что нейроны Tk-GAL42 ∩ Vglut демонстрируют повышение уровня кальция при дуновении (5d; видео №2).
Видео №2
В частности, сигнал GCaMP продолжал повышаться при многократном применении дуновений (красные временные окна (30–40 секунда и 50–60 секунда) на 5e). После прекращения воздействия дуновениями сигнал снижался до исходного уровня в течение 10 секунд (40-50 секунда на 5e). В отличие от дуновений воздуха, не удалось обнаружить изменений сигнала GCaMP в ответ на визуальный объект с предшествующими дуновениями или без них (синие временные окна на 5e).
Когда сигналы GcaMP были преобразованы в z- оценки и усреднены по временным окнам для дуновений или визуального объекта (5f), наблюдалось увеличение z-оценки примерно на 1.3 раза во время дуновений (слева на 5f), тогда как при демонстрации зрительного объекта такого увеличения не наблюдалось (справа на 5f). Следовательно, нейроны Tk-GAL42 ∩ Vglut кодируют дуновения (но не визуальный объект) в виде повышения уровня кальция.
Затем ученые решили установить, откуда информация о дуновениях передается нейронам Tk-GAL42 ∩ Vglut.
Поскольку ветер частично воспринимается сегментом a3 антенн 48, ученые предположили, что нарушение функции антенн может блокировать реакцию нейронов Tk-GAL42 ∩ Vglut на дуновения воздуха во время испытаний. Чтобы проверить эту гипотезу, функция a3 была специально нарушена либо хирургическим удалением, либо приклеиванием (5g). После этого была выполнена визуализация динамики кальция в нейронах Tk-GAL42 ∩ Vglut при воздействии дуновений.
Ожидаемо нарушение a3 заметно снижало повышение уровня кальция в нейронах Tk-GAL42 ∩ Vglut (5h). Дальнейшая количественная оценка показала, что нарушение a3 снижает максимальные сигналы GCaMP при воздействии дуновений примерно на 60% (5i).
Поскольку уже была установлена причинно-следственная связь между нейронами Tk-GAL42 ∩ Vglut и гейтированной зрительной аверсией (изображение №4), снижение реакции нейронов Tk-GAL42 ∩ Vglut на дуновения должно приводить к снижению гейтированной зрительной аверсии. В соответствии с этим рассуждением, хирургическое удаление сегментов а3 подавляло зрительное отвращение при воздействии дуновений (5j, 5k). Эти данные указывают на то, что дуновения активируют нейроны Tk-GAL42 ∩ Vglut через сегменты а3 антенн, вызывая зрительное отвращение.
Изображение №6
Пока что вышеописанные результаты опытов говорят об увеличении активности нейронов Tk-GAL42 ∩ Vglut на дуновения воздуха, но не визуальные объекты (5e, 5f). Следовательно, необходимо было установить, как тогда повышенная активность нейронов Tk-GAL42 ∩ Vglut транслируется в зрительное отвращение.
Как отмечают ученые, множество информации может кодироваться в гиппокампе (например, частота импульсов и временной паттерн импульсов). Зная это, ученые решили проверить, позволяют ли дуновения нейронам Tk-GAL42 ∩ Vglut кодировать визуальный объект как колебательную активность.
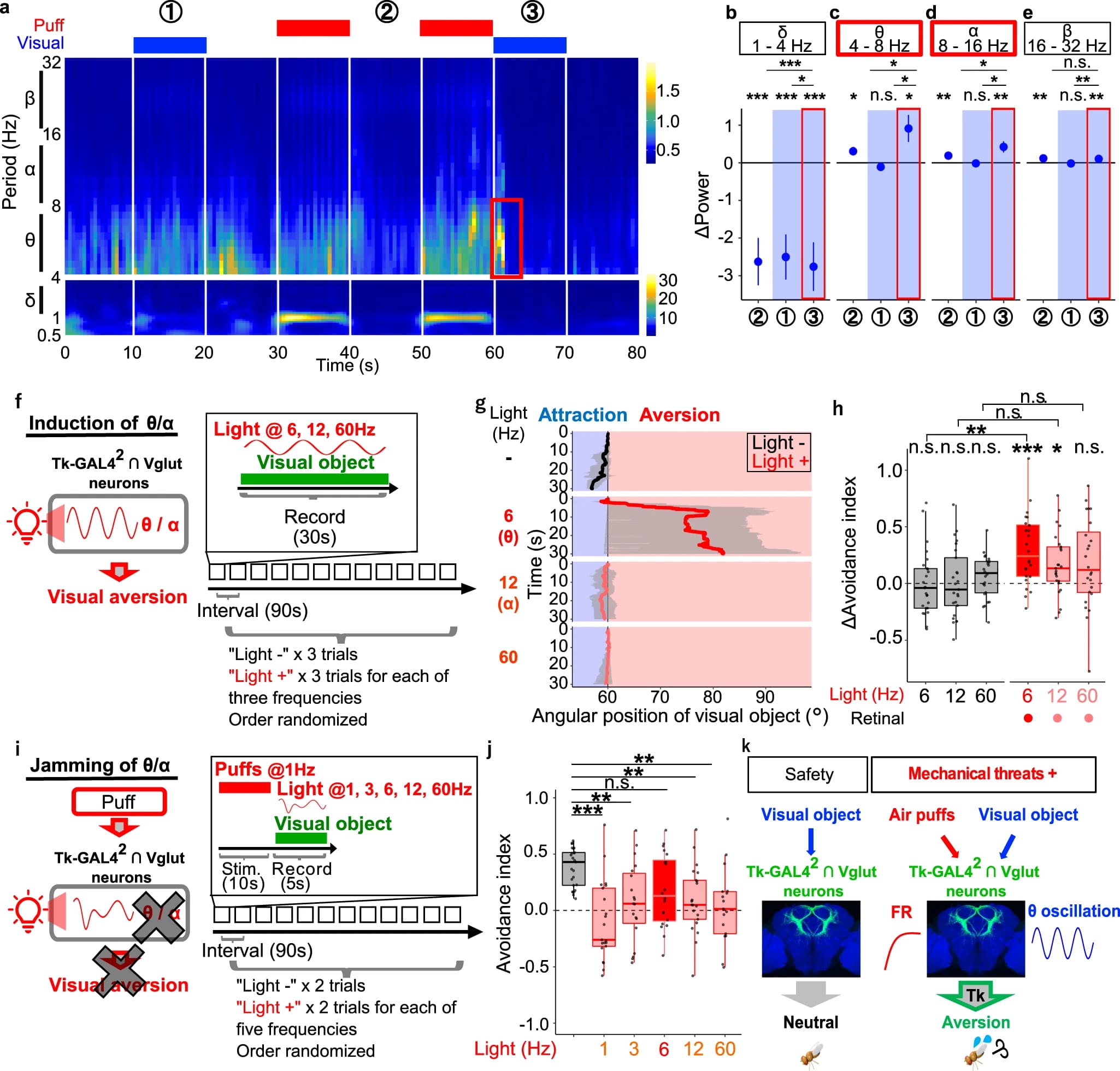
Сначала было подтверждено, что установка для визуализации кальция может обнаруживать колебательную активность как минимум до 25 Гц. Когда проводился анализ данных визуализации кальция (изображение №5), было обнаружено, что мощность колебаний θ (4–8 Гц) и α (8–16 Гц) временно увеличивалась, особенно когда зрительный объект предъявлялся после, но не до воздействия дуновений (6a).
Количественная оценка изменений мощности по сравнению с предыдущим интервалом времени 0.5 с (6b–6e) подтвердила, что колебания θ/α временно увеличивались, когда визуальный объект предъявлялся после дуновения (6c, 6d). Дуновения в чистом виде (без объекта) также увеличивали θ/α-колебания, но значительно в меньшей степени. С другой стороны, β-осцилляции незначительно усиливались в ответ на дуновение воздуха как таковое, а δ-осцилляции не проявляли стимульной специфичности (6b, 6e).
Поскольку колебания θ/α, но не δ/β, коррелируют со зрительным отвращением, эти данные позволяют предположить, что колебания θ и/или α могут быть причинно связаны со зрительным отвращением. Интересно, что усиление колебаний θ в ответ на визуальный объект было характерно для клеток, активированных дуновениями. Это наблюдение предполагает, что часть нейронов Tk-GAL42 ∩ Vglut, которая кодирует как дуновения, так и визуальный объект, может быть ответственна за зрительное отвращение, вызванное дуновением.
Затем ученые попытались проверить, влияют ли колебания θ/α в нейронах Tk-GAL42 ∩ Vglut на зрительное отвращение. Если это так, то фотоактивация нейронов Tk-GAL42 ∩ Vglut с частотами θ/α при предъявлении визуального объекта без предшествующих дуновений должна способствовать зрительному отвращению более эффективно, чем другие частоты (слева на 6f).
Ученые отмечают, что данный эксперимент отличается от того, что представлен на 3e-3h и изображении №4. В этом эксперименте ученые фотоактивировали нейроны Tk-GAL42 ∩ Vglut во время представления визуального объекта, чтобы имитировать колебания θ/α, которые возникают в ответ на визуальный объект.
В ходе опыта проводилась фотоактивация нейронов Tk-GAL42 ∩ Vglut с разной частотой, предъявляя при этом небольшой зрительный объект без предшествующих дуновений (справа на 6f). Несмотря на то, что общее количество доставленных фотонов оставалось постоянным на разных частотах, фотоактивация с частотой, соответствующей θ (6 Гц), наиболее эффективно стробировала визуальное отвращение (6g, 6h; видео №3). Напротив, фотоактивация нейронов Tk-GAL42 ∩ Vglut с другими частотами не вызывала зрительного отвращения на статистически значимых уровнях. Следовательно, тета (θ) с большей вероятностью, чем другие частоты, причинно способствует зрительному отвращению.
Видео №3
Значит ли это, что осцилляция θ необходима для зрительного отвращения в ответ на дуновения? Предыдущие исследования успешно подавляли колебательную активность, искусственно вызывая колебания нерелевантных частот. Таким образом, ученые хотели проверить, подавляет ли принудительная индукция частот, отличных от θ, во время визуального стимула визуальное отвращение в ответ на дуновения (слева на 6i). С этой целью на муху воздействовали дуновениями, а затем предъявляли визуальный объект, фотоиндуцируя различные частоты в нейронах Tk-GAL42 ∩ Vglut (справа на 6i).
Результаты опыта показали, что фотоиндукция всех частот, кроме θ (6 Гц), подавляла зрительное отвращение (6j). Таким образом, эти данные предполагают наличие причинно-следственной связи между визуально вызванными колебаниями θ и зрительным отвращением, вызываемым дуновениями.
Основываясь на полученных данных, ученые предложили следующий механизм, лежащий в основе зрительного отвращения к дуновениям воздуха (6k): без дуновений зрительная информация поступает в нейроны Tk-GAL42 ∩ Vglut, но не вызывает ответа; напротив, наличие дуновений вызывает повышение уровня кальция в нейронах Tk-GAL42 ∩ Vglut, позволяя им увеличивать θ-колебания при последующей визуальной информации, что, по крайней мере, частично способствует зрительному отвращению.

Изображение №7
Далее ученые решили выяснить, являются ли стробирование колебаний θ и возникающее в результате визуальное отвращение специфическими для небольшого объекта или вместо этого индуцируются также и другими визуальными объектами. Мухи обладают необычной особенностью — их привлекают темные вертикальные полосы (так называемая «фиксация на полосе»).
Таким образом, ученые исследовали, влияют ли потоки воздуха на фиксацию мух на полосе, используя тот же эксперимент, что и ранее, только вместо малого объекта были большие темные полосы (7a). Как и ожидалось, мухи фиксировались на полосе перед тем, как на них воздействовали дуновениями воздуха (черная линия на 7b; видео №4). Индекс фиксации, который является обратным индексу избегания, также показал сильное притяжение с дуновениями или без них (7c). Это наблюдение позволяет предположить, что, в отличие от реакции на небольшой объект, потоки воздуха не влияют на фиксацию на полосе.
Видео №4
Учитывая причинно-следственную связь между θ-колебанием и визуальным отвращением, можно предположить, что вертикальная полоса не вызовет θ-колебания. Чтобы проверить эту гипотезу, был проведен эксперимент по визуализации кальция, но с вертикальной полосой вместо небольшого объекта. Этот эксперимент показал, что вертикальная полоса действительно не вызывает колебания θ даже после воздействия дуновений (7d). Количественная оценка изменений мощности по сравнению с предыдущим временным интервалом 0.5 с подтвердила, что колебания θ не показали большего увеличения, когда вертикальная полоса была представлена после дуновений по сравнению с тем, когда были представлены только вертикальная полоса или только дуновения (7e).
Таким образом, установив, что стробирование θ-колебаний и возникающее в результате визуальное отвращение происходят в ответ на небольшой объект, а не на вертикальную полосу, ученые решили проверить, может ли принудительная индукция θ-колебаний отрицательно повлиять на фиксацию на полосе (слева на 7f). С этой целью колебания θ были оптогенетически индуцированы во время демонстрации вертикальной полосы, а поведение мухи записывалось (справа на 7f).
Фотоактивация с частотой, соответствующей θ, оказывала отрицательное смещение на фиксацию на полосе в тестовой группе (7g, 7h), в то время как частоты, отличные от θ (6 Гц), в той же группе не делали этого на статистически значимом уровне. В совокупности наши данные свидетельствуют о том, что даже фиксация на полосе, на которую не влияют дуновения, может быть аверсивно смещена искусственной θ-активностью в нейронах Tk-GAL42 ∩ Vglut.
В целом, данное исследование показывает, что дрозофилы в условиях опасности проявляют визуальное отвращение через Tk и одиночный кластер Tk-экспрессирующих нейронов. Данные показывают, как механические угрозы и визуальный объект могут быть закодированы в одном и том же наборе нейронов для адаптации зрительной реакции организма к опасной ситуации.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня труде ученые описали нейронный механизм, с помощью которого страх регулирует зрительное отвращение в мозгу дрозофилы. Кластер из 20-30 нейронов регулирует поведение мухи в ситуациях, которые она считает опасными.
В ходе опытов на мух воздействовали потоками воздуха, чтобы имитировать потенциальную опасность. Скорость движения особей после дуновений воздуха значительно увеличивалась. Затем в тестовую камеру помещали маленький черный объект. Сам он никак не влиял на поведение мух. Однако демонстрация этого объекта вслед за дуновениями воздуха вызывала у мух реакцию избегания визуального с ним контакта. Проще говоря, мухи старались не смотреть на объект, разворачиваясь к нему спиной.
Благодаря опытам ученые смогли установить нейроны, которые связаны с подобным поведением. Но вопросы еще остаются. Пока ученые не знают откуда эти нейроны берут входные данные и куда их передают, чтобы реакция избегания зрительного контакта реализовывалась. Ученые намерены продолжить свое исследование, сосредоточив внимание на построение нейронной карты, показывающей то, как именно страх влияет на зрение. По их мнению, такие исследования могут в будущем помочь в борьбе с психическими расстройствами, возникающими из-за преувеличенного страха, к которым можно отнести фобии и тревожные расстройства.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?