

Ученые провели партию в крестики-нолики с помощью ДНК. Звучит забавно, но это лишь верхушка айсберга притрушенная легким юмором от самих ученых. На самом же деле сегодняшнее исследование раскрывает новые методики контроля кинетики взаимодействия ДНК-наноструктур, которые в совокупности смогут выполнять сложные задачи. Использование ДНК как основных деталей системы — задача не из простых, но это не останавливает ученых, чьи умы полны идей, а сердца энтузиазма. И так, как же им удалось перемещать ДНК-блоки по заданному паттерну, что показало исследование и какие перспективы данной методики в будущем? За ответами обратимся к докладу исследовательской группы. Поехали.
Основа исследования
В данном исследовании ученые продемонстрировали новую методику перемещения ДНК плиток, в основе которой лежат принципы опорного соединения и смещения ответвления. Эти принципы схожи со стандартным смещением нитей ДНК, но нацелены на более крупные объекты, точнее на ДНК-структуры.
Данная методика позволила получить контроль над кинетикой ДНК плиток и реализовать желаемую цепочку реакций внутри мультиструктурных систем. Игра в крестики-нолики была проведена не забавы ради, а для демонстрации возможности смещения ДНК плитки в любом порядке и положении.
Для понимания механизмов биологических систем необходимо рассмотреть ее основные составляющие, в числе которых одними из важнейших являются молекулярные структуры. ДНК, как молекула-носитель информации, считается одним из лучших материалов для изучения и создания самоорганизованных наноструктур.
Среди методик, в основе которых лежит ДНК, больше и больше популярности последние годы приобретает методика ДНК-оригами, позволяющая создавать структуры из ДНК любой желаемой формы и конфигурации.
Составные элементы одного ДНК-оригами можно крутить-вертеть как угодно, но пока еще не удавалось достичь контролируемого взаимодействия между несколькими ДНК-оригами, как составными элементами большей, а потому и более сложной, структуры. На данный момент можно было только соединить/разъединить несколько ДНК-оригами.
В сегодняшнем исследовании ученые продемонстрировали новую методику взаимодействия ДНК-оригами. Имеется структура, в которую внедряется плитка ДНК-оригами, смещающая другую плитку из общего массива и соединяющаяся со структурой за счет связующего домена* на краю плитки (точка опоры/связи).
Связующий/связывающий домен* — домен белка, являющийся частью белковой цепи, но способный функционировать отдельно от нее.В качестве примера ученые создали три варианта реконфигурации: конкурентная, последовательная и кооперативная. Совокупность этих «движений» (пошаговая реконфигурация) была продемонстрирована как игра в крестики-нолики, когда у каждого игрока было 9 уникальных ДНК-оригами, которые можно перемещать в любом порядке на поле в 264х264 нм.
Формирование ДНК плиток
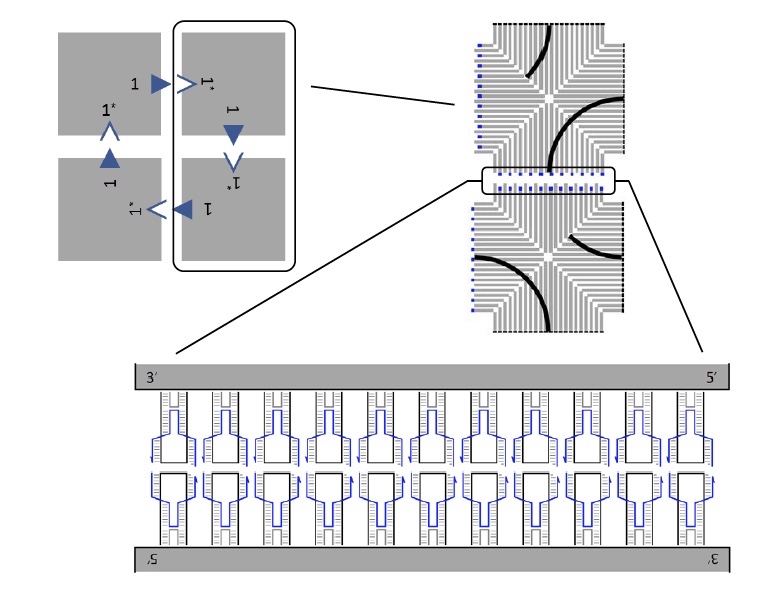
Структуры плиток ДНК.
Ранее, перед проведением данного исследования, ученые разработали методику создания индивидуальных ДНК плиток. У каждой плитки на двух краях имеется 11 скоб, каждая из которых участвует в стекинге.
На изображении выше показан массив из 4 плиток (сверху слева), каждая из которых соединяется с соседней посредством скоб.
Исследователи рассматривали несколько возможных вариантов создания более крупных массивов с большим числом плиток. Для этого, в теории, необходимо было, чтоб связь между плитками была достаточно слаба. Это позволило бы ее разорвать и перестроить массив. Однако наблюдения показали, что есть возможность создавать реконфигурируемые многоплиточные массивы даже при температурах, когда связь между плитками должна быть перманентной.
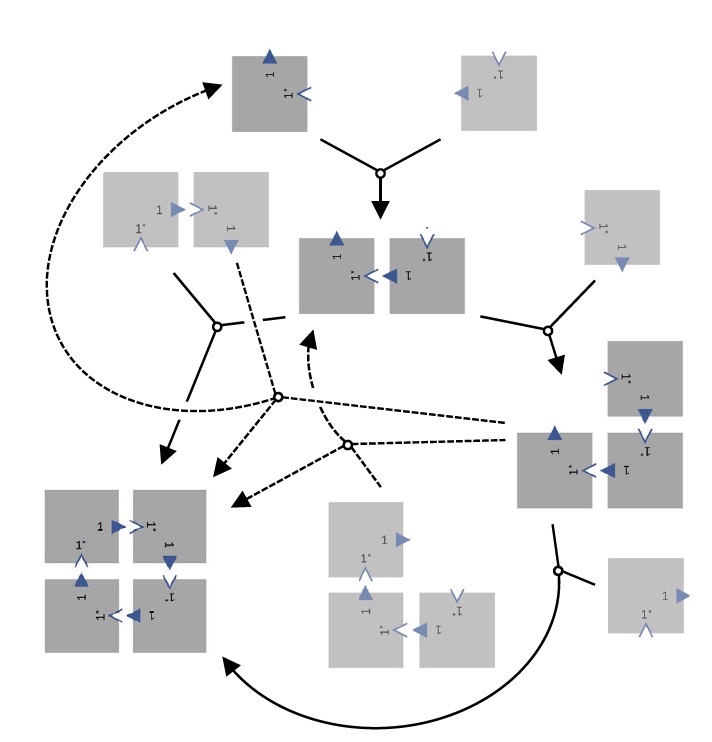
Реконфигурация плиток.
Это явление имеет два возможных пояснения. Первое — формирование связей происходит при достаточно высокой температуре, при этом попадая в температурный диапазон обратимости процесса. Второе — димер и тример подвергаются реакции вытеснения, посему образуется массив 2х2 при высвобождении мономера. Также могут делать и 2 копии тримеров, образующие массив 2х2 высвобождая димер.
Если же во время реакций смещения не будет самопроизвольного отсоединения плиток, то не будет возникновения кинетических ловушек, за счет чего плитки будут собираться в конфигурацию 2х2.
Необходимо было проверить, будет ли одна ДНК-оригами вытеснять другую в общей структуре, при этом не связываясь самопроизвольно внутри самой структуры. Другими словами, будут ли ДНК-оригами проявлять самодеятельность или нет. Для этого было проведено два эксперимента.

Практический опыт: 2 ДНК плитки (квадраты) + 1 ДНК треугольник, которые должны сформировать общую структуру.
В первом опыте участвовало 2 ДНК плитки и 1 ДНК треугольник, сам же эксперимент проводился при константной температуре. Структурно треугольник мог иметь либо такой же связующий домен, как и квадраты, либо обладал дополнительным связующим доменом, дополняющим соседний квадрат.
В первом варианте опыта 2 квадрата остаются связанными друг с другом, в то время как треугольник соединяется с одним из квадратов. Во втором варианте треугольник заменял один из квадратов, так сказать вытесняя его. Это на практике показало, что существует возможность изменять сложные ДНК-оригами. Но теперь нужно сделать так, чтобы этот процесс был подконтролен ученым, а не происходил сам по себе за счет естественных сил.
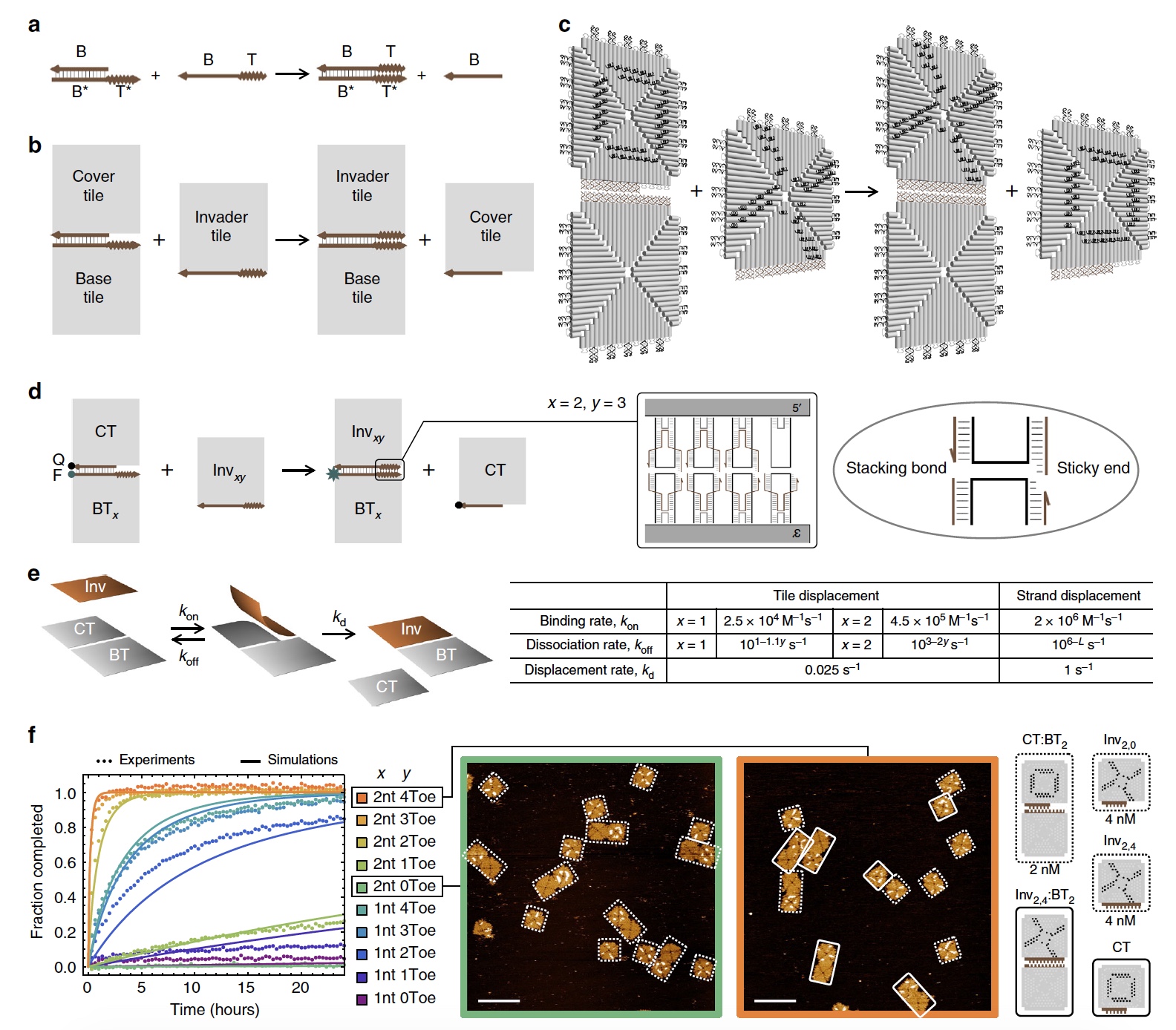
Изображение №1
Если говорить о реакциях смещения у цепей ДНК, то тут мы имеем цепь со связующим доменом, который соединяется со «свободным» комплементарным доменом. Так мы получаем двухцепочечную структуру, в которой происходит процесс миграции ветвей, когда цепи перемещаются через точку соединения (1а). Подобный процесс протекает и в реакциях смещения ДНК плиток (1b и 1с).
Из изображение 1с мы также может увидеть, что каждая ДНК плитка состоит из 4 равнобедренных треугольников, соединенными между собой скобами, о которых мы уже с вами знаем.
Касательно игры в крестики-нолики: для более удобной визуализации процесс смещения и реконфигурации участники были помечены крестиком (Х) и ноликом (О) посредством скоб двух разных цветов. Присмотритесь к изображению №1, на плитках видны Х (черный) и О (белый).
Кинетика реакций смещения ДНК плиток
Простыми словами говоря, достижение контроля над кинетикой процесса необходимо делать одни реакции более быстрыми, а другие более медленными. Таким образом можно достичь соединения, разъединения и реконфигурации плиток. Но нужно определить диапазон кинетики, позволяющей это осуществить. Для этого ученые создали точки соединения (между плитками) с разной силой, так сказать, сцепки.
Число скоб на соединительной в домене миграции ветвей было одинаковым для всех плиток. Но вот область доменов опорных (связующих) точек варьировалась от 0 до 4 (1d).
В эксперименте участвовала основная плитка («активны» все 4 точки связи) и придаточные плитки (число точек связи варьировалось от 0 до 4). В конце домена миграции ветвей 2 пары скоб были модифицированы флуорофором и гасителем. Если придаточная плитка сохраняет связь с основной, то флуорофор будет гаситься, что приведет к низкому флуоресцентному сигналу. А вот если придаточная плитка (с гасителем) будет отсоединена, флуоресцентный сигнал усилится. Таким образом можно будет определить как протекает процесс.
Спустя сутки можно было делать первые выводы из наблюдений. Ученые отмечают, что флуоресцентные траектории придаточных плиток с 0 соединительных скоб оставались практически неизменны. Также было замечено, что плитки с идентичной длиной соединительного ребра, но с большим число точек соединения, проявляют куда более быструю кинетику. Любопытно, что точки связи со скобами с 1-нуклеотидом и 2-нуклеотидом показывают очень разную скорость сатурации (насыщения).
Изображение 1е показывает математическую модель, использованную для численной оценки кинетики. Сами ученые говорят, что модель весьма проста и часто используется для оценки кинетики в реакциях смещения цепей ДНК, потому подходит и для плиток ДНК.
Сравнительный анализ (1f) симуляций (смоделированных реакций) и экспериментов дал возможность определить те параметры, которые влияют на процесс рекомбинации ДНК плиток.
Прежде всего ученые отмечают, что скорость соединения плиток в 10-100 раз (зависит от длины ребра соединения плитки) меньше, чем скорость соединения нитей ДНК. Общим между процессами смещения плиток и нитей является то, что с увеличением числа нуклеотидов в точках связи, экспоненциально уменьшается скорость распада. При этом у плиток эта скорость в 40 раз меньше, чем у нитей ДНК. Максимальный общий показатель смещения для ДНК плиток (скорость реакции) составил 4.5х105 М-1*с-1. Если концентрация низкая (<50 нM), то скорость бимолекулярного связывания будет ограничивать скорость смещения плиток. Если же концентрация больше 50 наномоль, то уже скорость мономолекулярного смещения будет ограничивать скорость смещения плиток.
Далее ученые провели несколько экспериментов, описывающих 3 основных типа реконфигурации: конкурентная, последовательная и кооперативная. А теперь давайте рассмотрим результаты опытов для каждой из них подробнее.
Конкурентная реконфигурация
Для достижения конкурентной реконфигурации ученые предложили использовать сигмоидальную функцию в ответ на концентрацию сигнала, таким образом получив систему реконфигурации на основе информации. Данная функция является важным элементом цифровых логических вычислений процессов смещения нитей ДНК, посему ее можно применить и для плиток ДНК тоже.
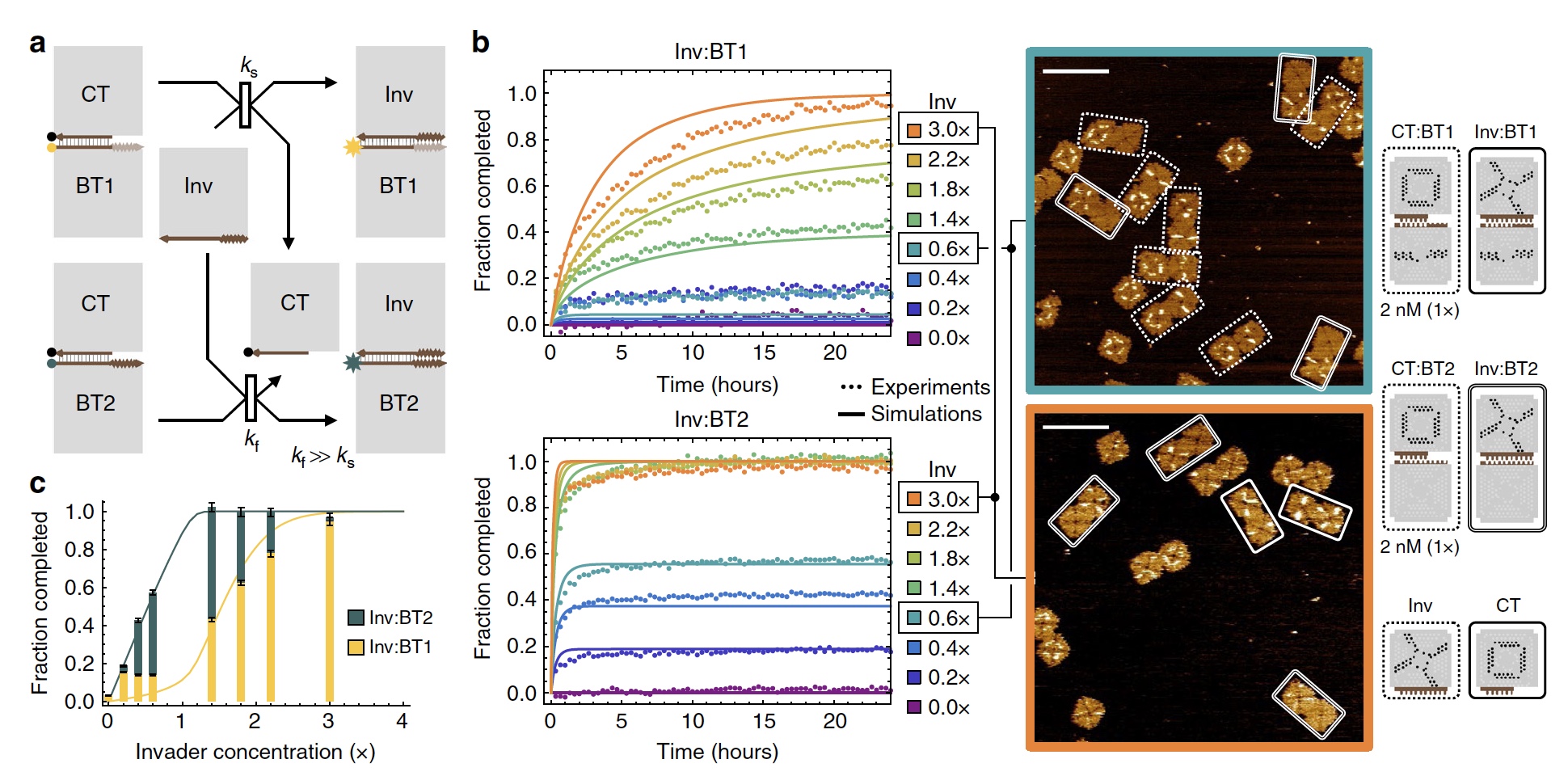
Изображение №2
Дабы продемонстрировать «работу» сигмоидальной функции ученые создали 2 конкурирующие реакции смещения плиток, которые активируются одной и той же придаточной плиткой. Разница в их скорости (2а).
У придаточных плиток точка связи это четыре 2-нуклеотидные скобы. Ожидается, что она будет соединяться с подходящей точкой связи и с четырьмя 1-нуклеотидными скобами. Скорость реакции же у придаточных плиток отличается в 18 раз. Когда у придаточной плитки концентрация была меньше 2 нМ, запускалась более быстрая реакция (2b). Если же концентрация была больше 2 нМ, то запускалась более медленная реакция.
Спустя 24 часа было установлено, что результат более быстрой реакции увеличивался линейно, а результат более медленной реакции показал сигмоидальную функцию (2с).
Последовательная реконфигурация
Следом за конкурентной реконфигурацией была последовательная. Данный тип куда сложнее, но дает больше контроля над процессом, если конечно все получится.
Итак, ученые создали массив 2х2, в котором первая придаточная плитка смещает другую плитку, освобождая ранее используемую точку связи. Далее уже вторая придаточная структура из 2 плиток смещает 2 плитки из массива. Своего рода каскадное смещение (3а).
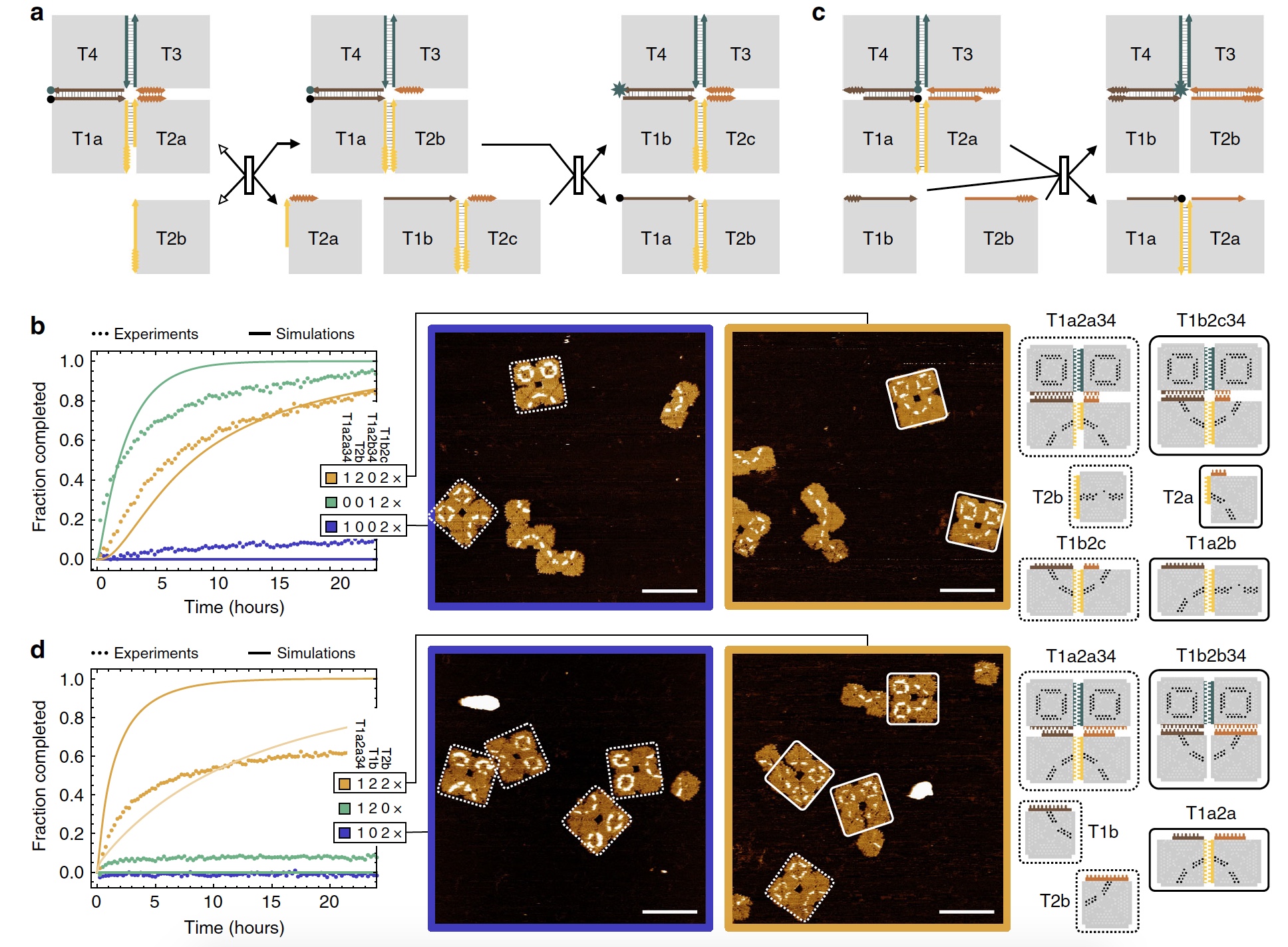
Изображение №3
Ученые подметили, что смещение по углам плиток проходит медленно, примерно в 100 раз медленнее, чем на остальном участке ребра плитки. Посему такие промежуточные состояния смещения необходимо учитывать для проведения более сложных процедур реконфигурации.
На изображении 3b мы можем увидеть вполне успешные результаты опыта с последовательной реконфигурацией (оранжевая кривая).
А сейчас еще немного креатива от исследователей. Обратите внимание на изображения выше (3b): массивы, которые реагировали на первую придаточную плитку, но не на вторую, в итоге выглядят как «грустная мордашка». А вот массивы, у которых связь с обоими придаточными плитками прошла успешно, выглядят как «улыбающаяся мордашка». А вы говорите, у ученых нет чувства юмора.
По словам ученых, чем сложнее структура участников реакции, тем сложнее сам процесс реконфигурации, но и тем интереснее ее результат. Использование двух типов участников (массива 2х2 и двухплиточной структуры) показало следующие результаты по эффективности смещения спустя 48 часов наблюдений: 83.3± 9.8% и 90.5 ± 6.1%. Другими словами, реакции смещения проходят отлично. Есть некие погрешности, но они незначительны.
Эксперименты с последовательной реконфигурацией позволили не только понять основные принципы последовательности точек связи и «каскадность» реакции, но и продемонстрировали новые особенности. Система способна реагировать на более чем 1 сигнал, указывающий какие инструкции требуют исполнения и что для этого имеется. Этот сигнал представлен в эксперименте в виде двух типов придаточных структур. Даже если первая придаточная структура входит в контакт с основой раньше второй, структура все равно показывает ожидаемую реконфигурацию.
Эти процессы по большей части протекают сами по себе. Ученые лишь слегка контролируют их, но не вмешиваются по полной программе. Следующим шагом в экспериментах является увеличение контроля над процессом реконфигурации и изучение возможности программирования этого процесса.
Кооперативная реконфигурация
В этом эксперименте было 2 придаточные плитки, которые соединялись с двумя краями массива 2х2 (3с). Если присутствует только одна плитка, то она должна переместится к центру массива. При этом отсутствие второй плитки позволяет обратить процесс, тем самым повторно отсоединив придаточную плитку от массива. При наличии обоих придаточных плиток возникает сразу два процесса миграции ветвей, которые сходятся в центре массива, а в результате мы получаем кооперативное смещение.
Опыт с флуоресценцией показал, что при наличии только одной придаточной плитки, флуоресцентный сигнал очень слаб, но усиливается если присутствуют обе плитки (3d: синяя и зеленая кривая — слабый сигнал, желтая — сильный сигнал).
Так же, как и в предыдущем эксперименте, массивы, которые взаимодействуют только с 1 придаточной плиткой, помечались как «грустная мордашка», а массивы и обе плитки — «улыбающаяся мордашка». При этом последние формировались в 68.0 ± 7.7% случаев.
Крестики-нолики
И, наконец, эксперимент «крестики-нолики». В данном опыте был использован массив 3х3, что позволяет провести 9 уникальных реакции смещения плиток в любом порядке (4а).
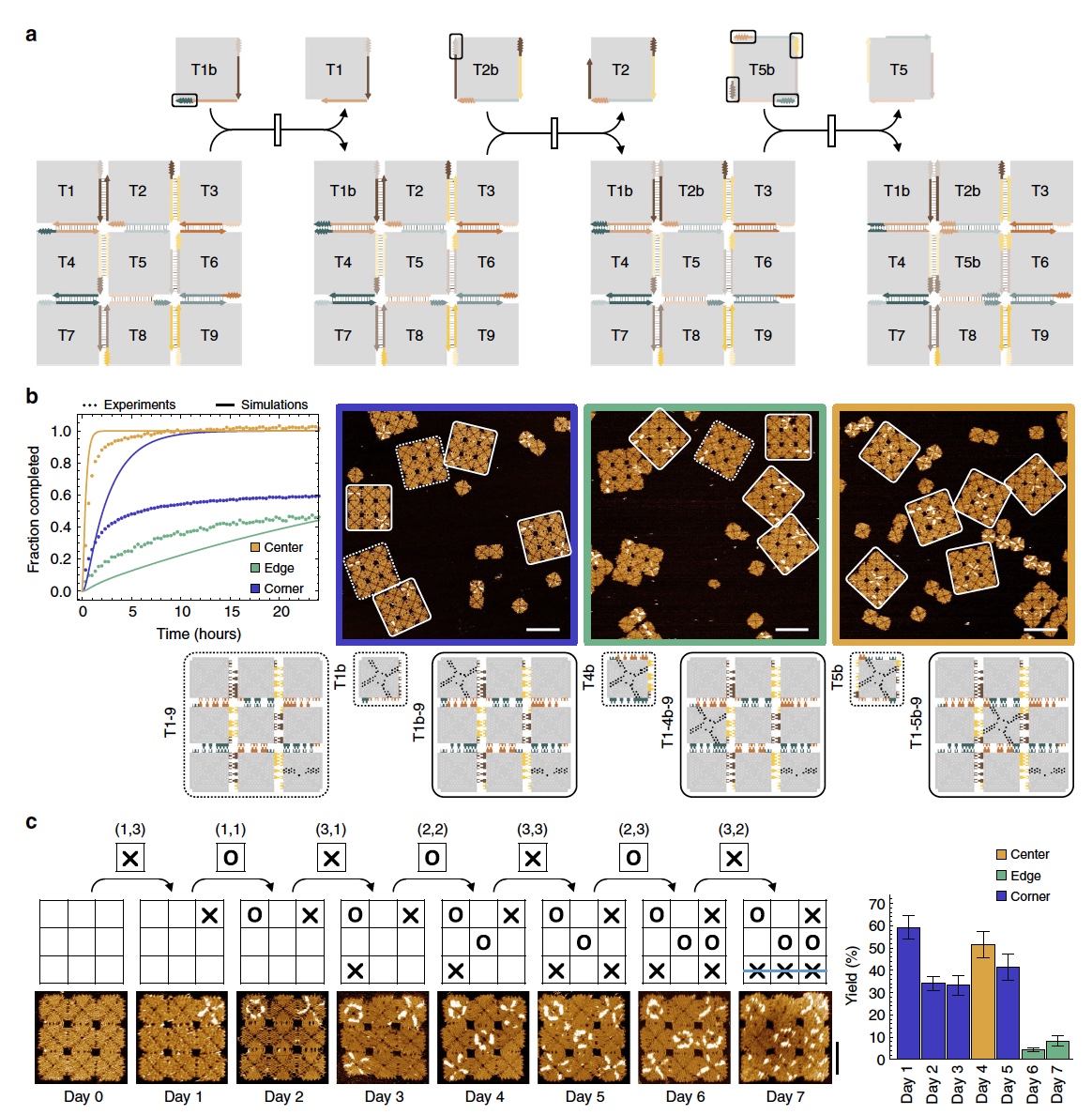
Изображение №4
Данный эксперимент является совокупностью сразу нескольких типов реакций: смещение угловой плитки, смещение краевой плитки и смещение центральной. По периметру массива между каждыми соседствующими плитками было по одной точке связи, всего 8 таких точек. 4 из них участвовали в смещении угловой плитки, когда придаточная плитка с соответствующей точкой связи контактирует с краевой плиткой рядом с угловой. В этот момент ветвь мигрирует внутри края плитки и через угол в 90°, а ранее соединенная с массивом угловая плитка высвобождается и интегрируется в сам массив (внутри массива, когда ранее была на краю). Другие 4 точки связи использовались для активации смещения краевых плиток.
Самым сложным в данном эксперименте было инициировать смещение центральной плитки, ввиду ограниченного числа доступных точек связи. Посему было добавлено 4 дополнительные точки между центральной плиткой и всеми соседствующими с ней (4а).
Наблюдения показали отличный результат. Все придаточные плитки были внедрены в массив, который перестраивался так, как и планировали ученые. Процент преобразования внутри массива был следующий: 78,4 ± 6,0% для смещения угловых плиток, 52,8 ± 6,0% для смещения краевых плиток и 100% — центральных.
А теперь перейдем непосредственно к самой игре, которая объединила в себе все типы реакции в виде каскада. Игровая доска представляла собой массив 3х3, все плитки были «чисты», то есть никак не помечены.
Каждый из игроков, коих было 2 естественно, получал в свое распоряжение 9 придаточных плиток, помеченных «Х» и «О». Совершение хода в данной игре это добавление в тестовую камеру этой плитки с интервалом в 24 часа (длинная партия в крестики-нолики, не правда ли).
Видео-демонстрация игры в крестики-нолики с помощью ДНК плиток.
Наблюдения (4b и 4с) показали, что игровое поле реагировало на все ходы, то есть реконфигурация протекала в соответствии с инструкциями со стороны игроков (добавление помеченных плиток).
Вся проблема была в том, что к концу игры на поле было очень много придаточных плиток, соответственно, увеличивалась и вероятность их самопроизвольного смещения. Таким образом точность выполнения реконфигурации также снижалась. К концу игры этот показатель был всего лишь 8.3 ± 2.3%. Однако результата добиться все же удалось, как мы можем видеть с изображений выше.
Для более детального ознакомления с нюансами исследования и экспериментов настоятельно рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Игра в крестики-нолики ДНК плитками была проведена не ради забавы, а как метод демонстрации весьма серьезного исследования, раскрывающего возможность манипулировать процессами смещения элементов в наноструктурах, построенных из ДНК.
Конечно, многое еще предстоит отшлифовать: число придаточных плиток, влияющих на конечный показатель реконфигурации, скорость протекания реакций, точность смещения, расширение возможных вариантов смещения и, конечно же, увеличения габаритов реконфигурируемой ДНК структуры.
Сейчас многие ученые все больше и больше внимания уделяют весьма нестандартным и нетривиальным механизмам выполнения процессов, которые уже стали стандартными. Хранения и обработка информации, диагностика и лечение заболеваний, изучение сложных систем, в том числе и биологических — все эти процессы можно выполнять с помощью ДНК, поскольку этот невероятно малый объект скрывает в себе невероятные свойства и характеристики, которые нам еще предстоит раскрыть.
Кстати, оказывается если ввести в google запрос «крестики-нолики», то можно сыграть в эту игру. :) (не знаю для кого как, но я этого не замечал раньше)
И, конечно же, пятничный оффтоп (продолжу новогоднюю тематику, если вы не против):
Немного милоты под елку :)
Благодарю за внимание, оставайтесь любопытствующими и отличных вам выходных, ребята.
Немного милоты под елку :)
Благодарю за внимание, оставайтесь любопытствующими и отличных вам выходных, ребята.
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас оформив заказ или порекомендовав знакомым, 30% скидка для пользователей Хабра на уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2650 v4 (6 Cores) 10GB DDR4 240GB SSD 1Gbps от $20 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
VPS (KVM) E5-2650 v4 (6 Cores) 10GB DDR4 240GB SSD 1Gbps до весны бесплатно при оплате на срок от полугода, заказать можно тут.
Dell R730xd в 2 раза дешевле? Только у нас 2 х Intel Dodeca-Core Xeon E5-2650v4 128GB DDR4 6x480GB SSD 1Gbps 100 ТВ от $249 в Нидерландах и США! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?