

Одной из основных отличительных черт любого живого организма является способность к сохранению и воспроизведению необходимой информации для создания себе подобных. В первую очередь это проявляется в репликации ДНК, когда из одной родительской ДНК появляется пара дочерних, являющихся точной копией своего прародителя. В природе этот процесс наблюдается повсеместно, однако воссоздать его в лабораторных условиях с нуля крайне сложно, тем не менее, вполне реально.
Ученые из института биохимии им. Макса Планка (Германия) успешно создали биологическую систему, которая обладает способностью к репликации собственного ДНК. Какие методики были применены для создания синтетического реплицирующего ДНК, насколько эффективна полученная система и что данное открытие значит для современной синтетической биологии? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
В области создания искусственных биологических систем способность к репликации являются ключевым аспектом полноценности созданной системы. Достичь этого, по словам ученых, можно посредством реализации базисных составляющих: репликация*, транскрипция* и трансляция* ДНК.
Репликация ДНК* — процесс создания из одной материнской ДНК двух дочерних молекул ДНК.
Транскрипция* — процесс синтеза РНК с использованием ДНК в качестве матрицы, т.е. процесс переноса генетической информации с ДНК на РНК.
Трансляция* — процесс синтеза белка из аминокислот на матрице РНК, осуществляемый рибосомой.Другими словами, чтобы создать полноценную искусственную жизнь, необходимо реализовать самокодированное воспроизведение, т.е. саморепликацию. Это крайне сложный и запутанный процесс.
Авторы исследования отмечают, что в системах, основанных на существующей биохимии, например MPC (minimal protein-based cells), саморепликация требует полного переосмысления молекулярной биологии, включая репликацию, транскрипцию и трансляцию ДНК, с учетом полностью бесклеточной среды.
Синтез белка из ДНК может быть достигнут в определенных рекомбинантных системах на основе фаговых* РНК-полимераз — основных частей механизма трансляции Escherichia coli и системы минимальной регенерации энергии, т.е. PURE — Protein synthesis Using Recombinant Elements (синтез белка с использованием рекомбинантных элементов).
Бактериофаги или фаги* — вирусы, избирательно поражающие бактериальные клетки и клетки архей. В биологии применяется в качестве вектора (ДНК для передачи генетического материала внутрь клетки).Тем временем остается крайне сложным для воссоздания процесс ДНК репликация генома на базе транскрипции и трансляции (TTcDR — transcription–translation-coupled DNA replication), кодирующая все макромолекулярные компоненты системы PURE с помощью самокодируемой реплисомы*.
Реплисома* — мультибелковый комплекс, за счет которого проходит репликация ДНК бактерии.Помочь реализовать методику TTcDR может ДНК репликация на базе ДНК-полимераз (DNAP) из фагов (например, Phi29).
Однако в методике TTcDR достичь полноразмерного генома Phi29 можно было исключительно путем блокировки части реплицирующих факторов.
Все вышеописанные способы имеют свои теоретические преимущества, однако на практике они не давали полноценного результата. В рассматриваемом нами сегодня труде ученые описывают систему трансляции, которая обеспечивает самокодированную репликацию и экспрессию больших геномов ДНК в четко определенных бесклеточных условиях. В частности, была достигнута саморепликация генома размером более 116 т.п.н. (тысяч пар нуклеотидов), охватывающего полный набор факторов трансляции Escherichia coli (кишечная палочка), все три рибосомных РНК, систему регенерации энергии, а также РНК- и ДНК-полимеразы*.
Полимераза* — фермент, выполняющий синтез полимеров нуклеиновых кислот. ДНК-полимераза синтезирует ДНК, а РНК-полимераза синтезирует РНК. Данный процесс протекает за счет комплементарного копирования родительских ДНК или РНК.Помимо этого, созданная система продемонстрировала возможность синтезировать более 30 факторов трансляции, половина которых экспрессирует в количествах равных или превышающих вводные.
Результаты исследования
На первом этапе ученые протестировали самокодированный Phi29-DNAP-зависимый TTcDR, используя стандартный протокол системы PURExpress.
Кодирующую область Phi29-DNAP, фланкированную промотором* Т7, сначала клонировали в вектор pCR-Blunt TOPO (pREP, 1a).
Промотор* — последовательность нуклеотидов ДНК, используемая РНК-полимеразой в качестве области для начала транскрипции.
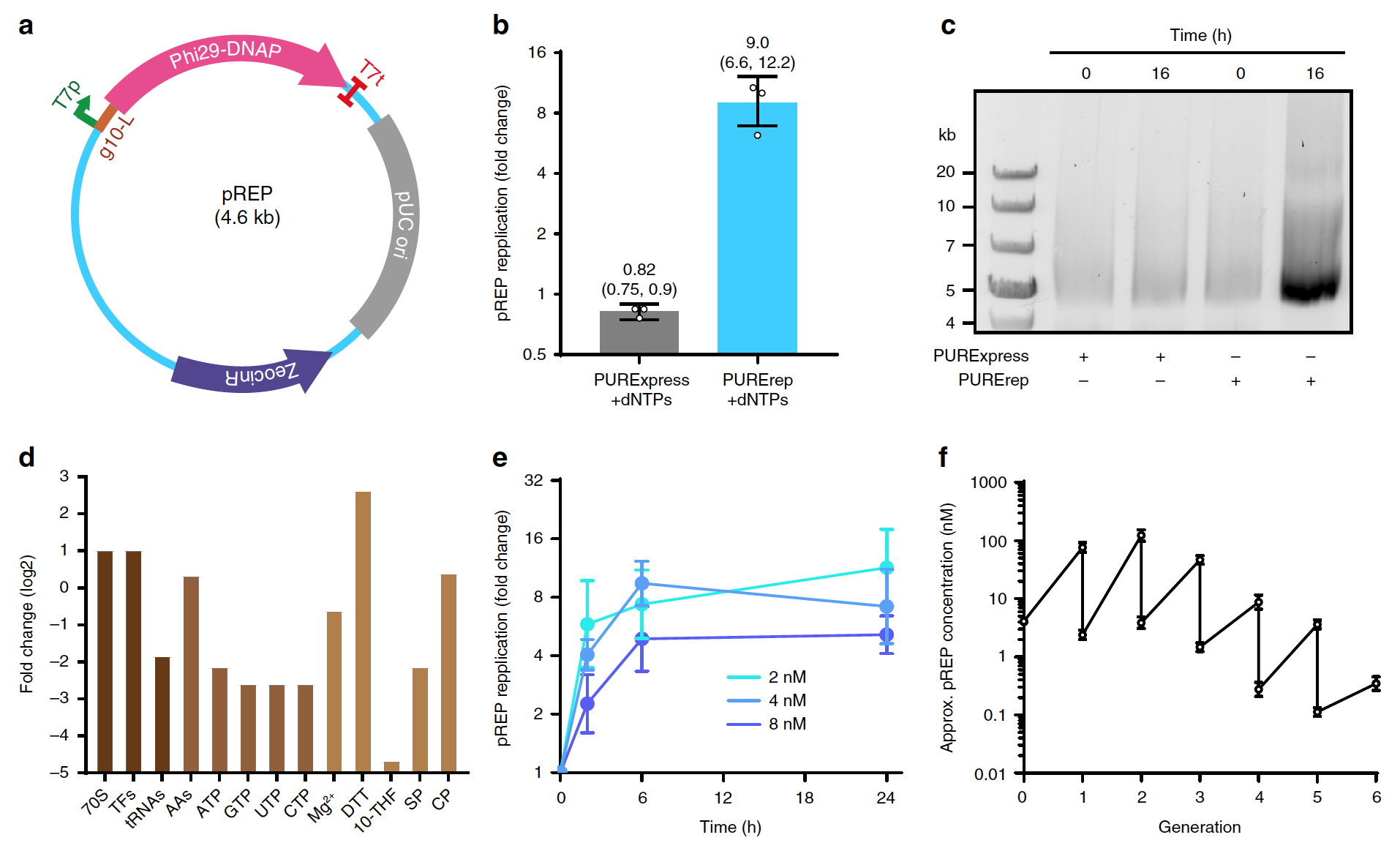
Изображение №1
В теории данная архитектура должна обеспечить спонтанную РНК репликация по типу катящегося кольца* с помощью самокодируемого DNAP без дополнительных белков репликации или предоставляемых извне ДНК-праймеров.
Репликация по типу катящегося кольца* — процесс однонаправленной репликации нуклеиновой кислоты, во время которого происходит быстрый синтез множественных копий кольцевых молекул ДНК или РНК.Однако, используя стандартную реакцию PURExpress с dNTP (нуклеотид, содержащий дезоксирибозу C5H10O4) и 4 нМ pREP (среда LB с добавлением зеоцина C55H86N20O21S2), обнаружить синтез ДНК не удалось ни с помощью электрофореза в агарозном геле*, ни с помощью полимеразной цепной реакции в реальном времени* (1b и 1c).
Электрофорез ДНК в агарозном геле* — метод разделения фрагментов ДНК по их длине, основанный на определении скорости движения фрагментов разной длины в момент движения в геле под действием внешнего электрического поля.
Полимеразная цепная реакция в реальном времени* — метод одновременного создания дополнительных копий участков ДНК и измерения количества данной молекулы ДНК.Чтобы улучшить репликацию ДНК без участия специализированных систем PURE, было решено оптимизировать стандартный протокол реакции PURExpress. Для этого было увеличено относительное количество факторов трансляции, рибосом и восстановителя при одновременном снижении уровней тРНК (транспортная РНК) и rNTP (рибонуклеозид трифосфат) (1d).
Используя этот оптимизированный состав PURE (PURErep), удалось добиться ?5–12-кратной репликации мономерных звеньев pREP в реакциях TTcDR (1b и 1e). Полноразмерный синтез pREP был подтвержден MluI-расщеплением* продукта репликации (1c).
MluI* является коммерчески доступной эндонуклеазой рестрикции, т.е. ферментом, катализирующих реакцию гидролиза нуклеиновых кислот.В качестве оценочного фактора активности трансляции выступила экспрессия зеленого флуоресцентного белка (sfGFP). Было выявлено, что изменение состава PURE привело к снижению синтеза белка на 20-40% по сравнению с PURE без TTcDR. Тем не менее, такое снижение экспрессии белков остается приемлемым, учитывая улучшение совместимость системы PURErep с репликацией ДНК.
Анализ репликации ДНК на основе qPCR выявил устойчивое время удвоения (1–2 часа) для различных начальных концентраций матрицы, при этом репликация ДНК продолжалась даже через 24 часа при 30 °С (1е).
TTcDR также был устойчив в течение более пяти поколений последовательного разбавления, когда лишь 4% готового продукта реакции PURErep/pREP непосредственно переносили в новую смесь PURErep (1f). Основным выводом данного наблюдения является то, что продукты TTcDR могут служить шаблонами для саморепликации ДНК в течение нескольких поколений.
Ученые отмечают, что для репликации по типу катящегося кольца характерно значительное количество продукта с низкой электрофоретической подвижностью, что и наблюдалось во время опытов. Эти продукты реакции могут быть представлены конкатемерами* с большой молекулярной массой и/или кластерами ДНК/MgPPi.
Конкатемер* — множественные копии ДНК последовательностей, собранные в последовательный кластер.Также было обнаружено формирование продуктов размером ~ 5 т.п.н. в необработанных образцах. Из этого следует, что реакции TTcDR могут давать значительные количества мономерных копий pREP.
Кроме того, ученым удалось трансформировать продукты E.coli после удаления родительской плазмы. Полученные очищенные продукты реакции были идентичны по размеру с мономерными pREP.
На следующем этапе ученые решили создать набор генов, кодирующих важные компоненты реакции PURE: 31 существенный фактор трансляции (ФТ) бактерии E. coli.
Для этого было проведено исследование TTcDR из pREP (4.6 т.п.н.) в сопряжении с каждой из трех больших плазмид*: pLD1 (30 т.п.н., 13 ФТ), pLD2 (20 т.п.н., 8 ФТ) и pLD3 (23 т.п.н., 9 ФТ). Все они были клонированы для обеспечения рекомбинантной экспрессии 30 из 31 факторов трансляции бактерии E. coli.
Плазмиды* — небольшие молекулы ДНК, способные к самостоятельной репликации.Продукты TTcDR всех четырех плазмид (включая pREP) демонстрировали идентичные особенности MluI-рестрикции: клональные плазмиды, обычно размножающиеся в E.coli (2a).
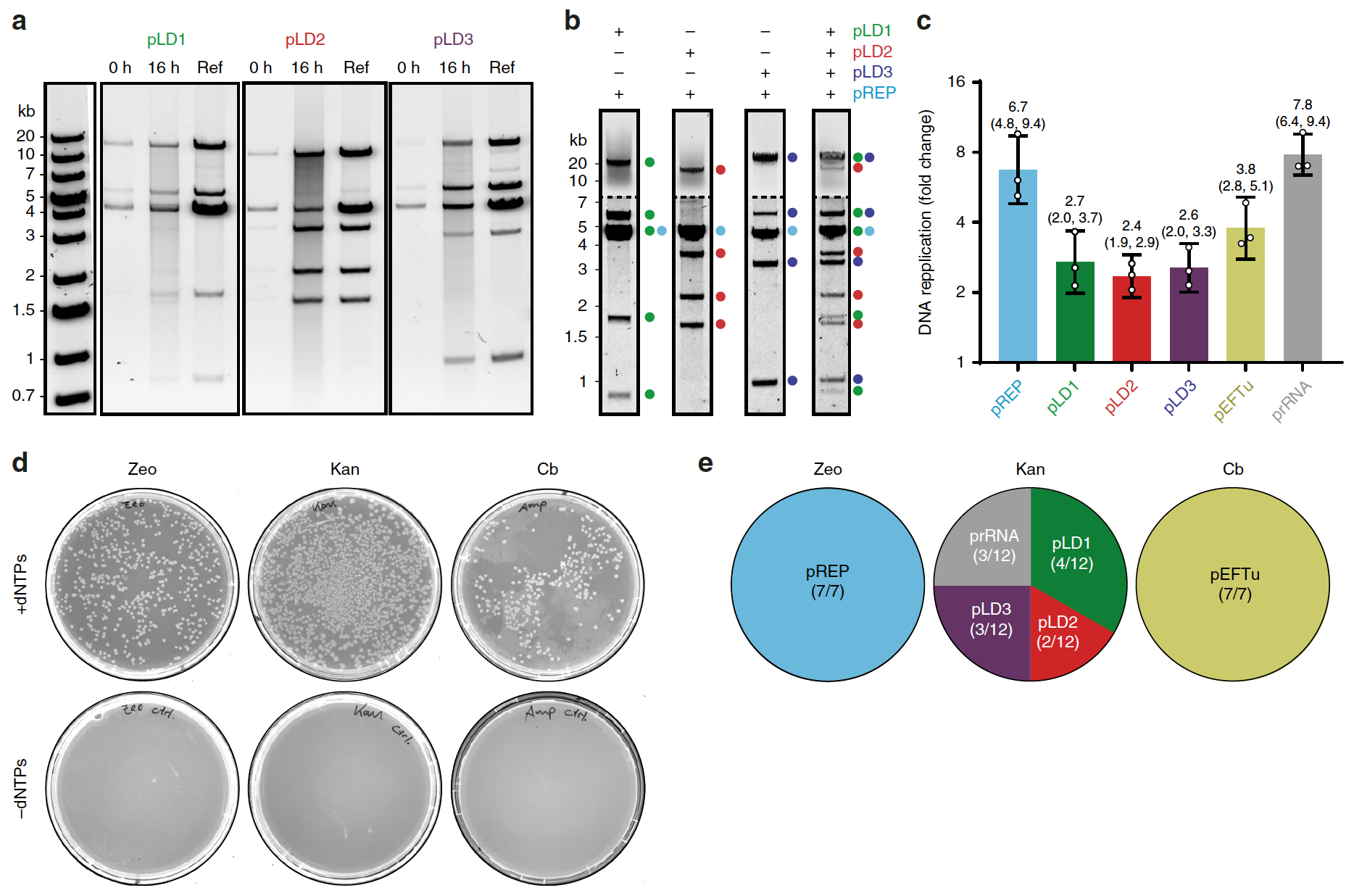
Изображение №2
Также стоит отметить, что продукты pLD TTcDR могут быть непосредственно трансформированы в E.coli, где они сохраняются в виде мономерных плазмид.
Модифицированная смесь PURErep также обеспечивала полную репликацию всех трех плазмид pLD вместе с PURErep во время реакции в общей среде (2b).
На этом ученые не остановились и уже на следующем этапе исследования попытались расширить генетическую нагрузку системы TTcDR путем совместной репликации плазмид, кодирующих дополнительные компоненты системы PURE: EF-Tu (pEFTu), которая отсутствует в системе pLD, а также рибосомальный РНК-оперон* rrnB (прРНК прекурсорная рРНК), которая кодирует 23S рРНК (рибосомная РНК), 16S рРНК, 6S рРНК и тРНК (транспортная РНК, в данном случае Glu2) (2c).
Оперон* — функциональная единица генома одноклеточных, в состав которой входят гены, кодирующие белки.Эксперименты с qPCR, нацеленные на плазмид-специфичные ампликоны*, подтвердили, что мономерные звенья всех шести плазмид (общая длина ДНК составила 93 т.п.н.) реплицировались примерно в 2-8 раз по сравнению с исходным числом (2c).
Ампликон* — внехромосомная единица амплификации (образования копий участков ДНК).Дополнительным подтверждением успешности совместной репликации плазмид стало образование колоний, устойчивых либо к зеоцину (pREP), либо к канамицину (плазмиды pLD и прРНК), либо к карбенициллину (pEFTu). Эти колонии стали результатом трансформации продуктов реакции PURErep, обработанных DpnI (2d).
Выбранные 26 клональных колоний с последующим анализом рестрикционных особенностей еще раз подтвердили успешность TTcDR всех шести плазмид (2e).
Используя тот же метод, что был описан ранее, ученые провели еще одну процедуру совместной репликации пяти дополнительных плазмид: гены системы минимальной регенерации нуклеозидтрифосфата на основе креатинкиназы (pCKM), аденилаткиназы (pAK1) и нуклеозиддифосфаткиназы (pNDK); а также T7-РНК-полимеразы (T7RNAP) и пирофосфатазы (pIPP).
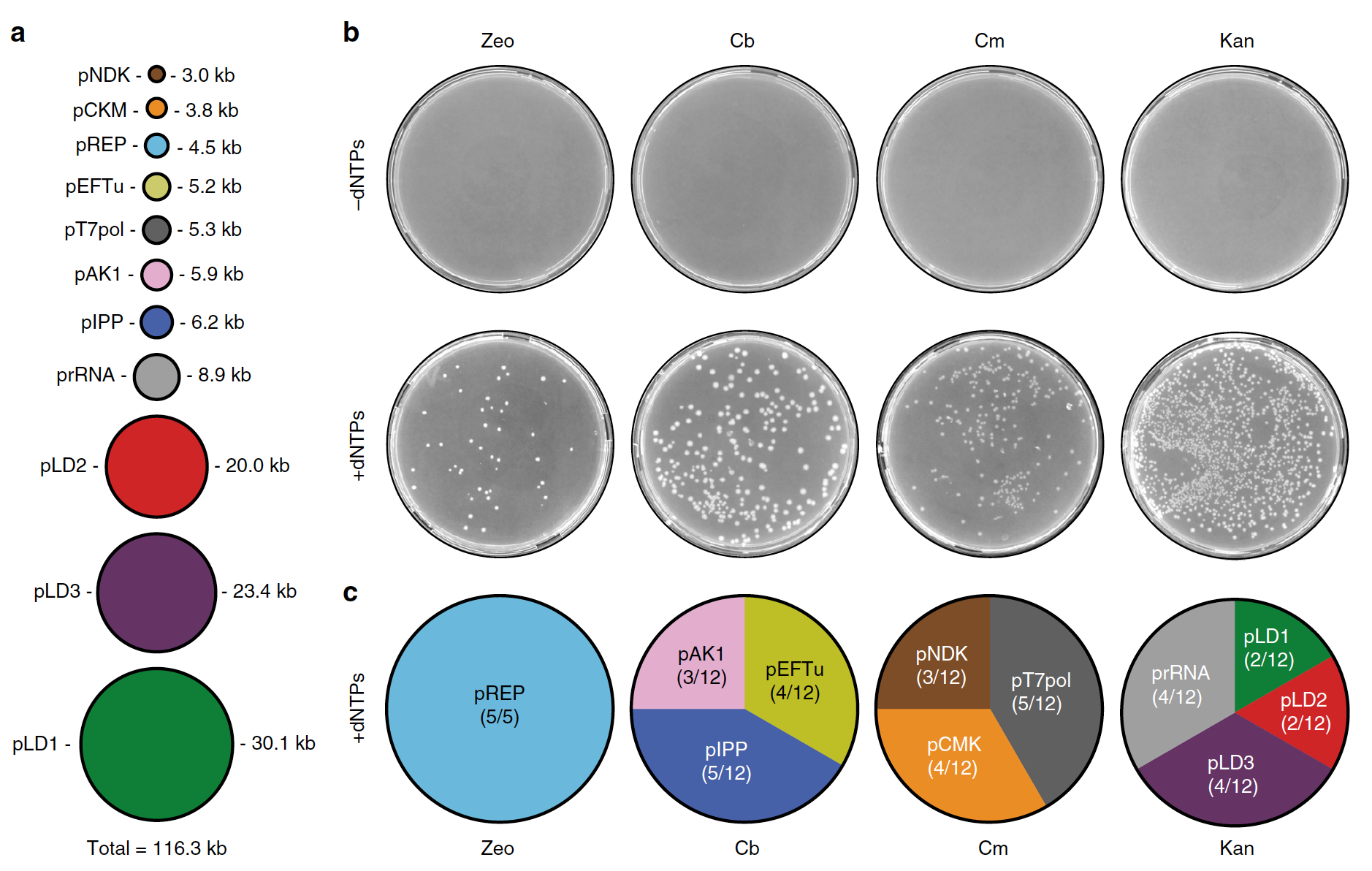
Изображение №3
При общем размере 116.3 т.п.н. данный набор из 11 плазмид достигает > 100% предполагаемой длины генома, предложенной для минимальной, самореплицирующейся системы, зависящей только от низкомолекулярных питательных веществ (3а).
Далее ученые решили проверить, может ли PURErep во время репликации обеспечивать параллельную экспрессию генов. Для этого была рассмотрена мультицистронная экспрессия ФТ, кодируемых на трех плазмидах pLD: pLD1, pLD2 и plD3 (не включая pEFTu).
Чтобы исследовать, способна ли PURErep в целом поддерживать мультицистронную экспрессию из этих плазмид, была проведена бесклеточная экспрессия из каждой отдельной плазмиды в присутствии BODIPY-Lys-tRNALys, что обеспечивает флуоресцентную маркировку продуктов трансляции.
Чтобы улучшить чувствительность обнаружения и дать возможность количественного определения вновь синтезированных белков, дополнительно был проведен количественный анализ экспрессии белка на основе масс-спектрометрии с использованием стабильных изотопных меток.
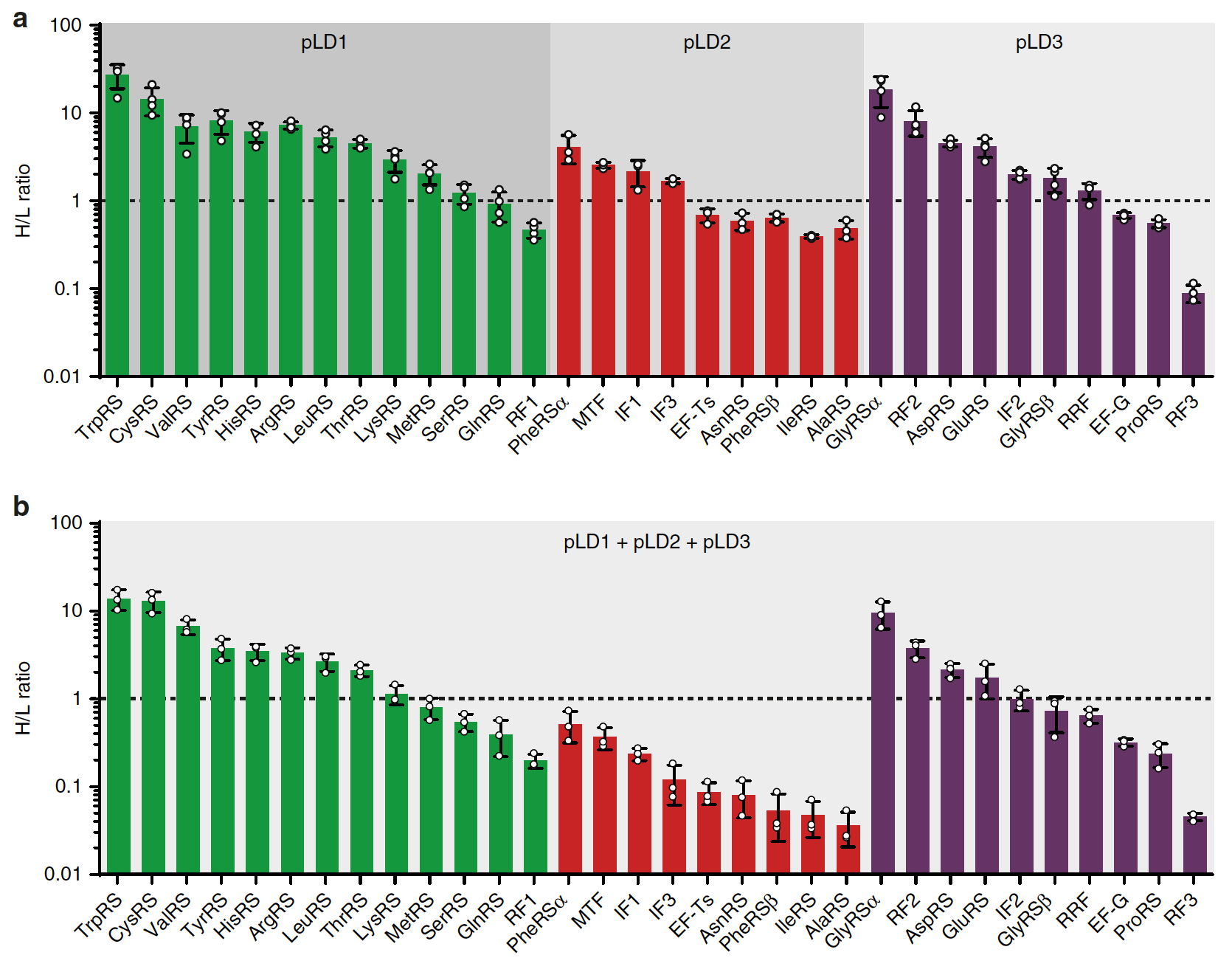
Изображение №4
Данный анализ предоставил убедительные доказательства синтеза всех субъединиц ФТ белков, кодируемых pLD (4а). Также наблюдалась частичная или полная регенерация белков, кодируемых как в pLD2, так и в pLD3.
Анализ экспрессии показал, что даже в не модифицированной PURErep в сочетании с pREP наблюдается полная репликация 32 ФТ цидронов, кодируемых pLD, и экспрессия примерно половины кодированных пептидных цепей ФТ, число которых соответствует или превышает исходное.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В данном труде ученым удалось достичь того, о чем многие их коллеги ранее могли только мечтать — создать искусственную систему, которая способна само-реплицировать.
Утрировано говоря, это означает, что им удалось сделать первые шаги к созданию системы, которая будет самовоспроизводиться, тем самым максимально имитируя биологические процессы.
Авторы исследования крайне довольны результатами и планируют в будущем расширить искусственный геном дополнительными сегментами ДНК. Они хотят создать систему с оболочкой, способную сохранять жизнеспособность, впитывая питательные вещества и избавляясь от отходов. Подобная искусственная структура может быть использована в качестве специализированного производственного био-устройства для природных веществ или в качестве основы для создания еще более сложных систем.
Клонирование — процесс сложный, в чем нет сомнений, однако создание полноценной, жизнеспособной синтетической системы — совсем другой уровень. Кто-то скажет, что подобные исследования опасны, ибо никто не вправе брать на себя обязанности матушки-природы. Тем не менее, любопытство человека практически невозможно унять. Наше желание все понять и все повторить является одним из движущих фактором прогресса науки. Хорошо это или плохо — вопрос, на который нет однозначного ответа. Нам остается лишь восхищаться достижениями светлых умов и с опаской смотреть в будущее, одновременно надеясь на утопию.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы :)
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


OleksiyT
E.coli, серьёзно?
Ну конечно, почему не выбрать для экспериментов по саморазмножению то, что чистый яд для человека.
Как будто Covid-19 мало.
Desiderio
Но ведь E.coli живут в каждом человеке. И только некоторые редкие их штаммы могут быть всего лишь вредными, но никак уж «чистый яд».
OleksiyT
Я в курсе, что сдавая анализы на воду проверяется уровень E.coli.
И допустимая норма для чистой воды: 0.
Т.е. их там вообще быть не должно, вы, как организм, не должны увеличивать их количество в себе.
И вот какие-то ребята решают именно их сделать саморазмножаемыми, а не что-то нейтральное.
Это, действительно, только мне кажется ненормальным?
Desiderio
Анализы «на воду» здесь не причём. Естественно, что чистая вода не должна содержать «ничего кроме воды», а тем более содержимого чьих-то кишечников.
Но для микробиологии E.coli это самая распространённая и самая изученная бактериальная культура. Лучше уж работать с тем, что уже давно и хорошо исследовано, чем создать какой-нибудь новый неизвестный аналог.
Jogger
Допустимая норма живых хомячков в чистой воде: 0
Количество живых хомячков в кишечнике человека: 0
Хомячки ГОРАЗДО опаснее чем E.coli! Даже один живой хомячок в кишечнике приведёт к серьёзным проблемам со здоровьем!
Вы должны быть рады, что учёные проводят тесты на безопасных E.coli, а не на смертельных хомячках.
ctc
DGN
Опыты проводились, пруф, на самом деле все не так плохо, а быть может, раз уж такие эксперименты проводятся, кому-то было даже очень хорошо…
dabar347
>И вот какие-то ребята решают именно их сделать саморазмножаемыми,
E.coli и без исследователей прекрасно размножаются
Jogger
Вы же в курсе что E.coli живут в вашем кишечнике постоянно и являются частью нормальной микрофлоры?
dmrnk
NaCl, серьезно?
Ну конечно, почему не выбрать для преправы к еде то, что чистый яд для человека. Как будто дигидрогена монооксида мало.
artemerschow
E.coli широко используется в микробиологии, биологической инженерии как модельный организм и для синтеза чужеродных белков. Например, весь человеческий инсулин синтезируется именно модицифированными E.coli
en.wikipedia.org/wiki/Escherichia_coli#Model_organism_in_life_science_research
en.wikipedia.org/wiki/Escherichia_coli_in_molecular_biology