

Без преувеличения мозг это одно из самых мощных оружий, коим может обладать живое существо. Быстрые лапы, острые когти, сильные мышцы и прочие физические атрибуты, конечно, тоже вносят свой вклад в борьбу за место под солнцем. Но именно интеллект, способность осмысленно оценивать обстановку, принимать взвешенные решения и планировать свои действия — эти навыки помогли многим видам дожить до наших дней куда сильнее, чем физическая сила и ловкость. Несмотря на всю важность мозга, который управляет всеми процессами в организме, словно центральный процессор в компьютере, о его происхождении мало что известно. Мы знаем, что существа с мозгами появились сотни миллионов лет тому назад, но вот как эти первобытные мозги возникли — остается загадкой. И вот ученые из Европейской молекулярно-биологической лаборатории выдвинули гипотезу о том, морская губка, у которой нет мозга, может быть ключом к секретам эволюции этого органа. Что у губки вместо мозгов, какие гены у губок указывают на эволюцию мозга, и причем тут их пищеварительная система? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Морских губок (Porifera на 1A) не назовешь сложными организмами, ибо у них нет полноценных нервных клеток, мышц или даже кишечника. Однако они все же являются многоклеточными. У губок имеется три основных типа клеток, которые в совокупности создают эффективный водяной насос для фильтрации пищи и удаления отходов (1B-1C).
Изображение №1
Хоаноциты (2-10 мкм в диаметре), снабженные жгутиками и микроворсинками, образуют хоанодерму, необходимую для фильтрации (1C).
Пинакоциты (20 мкм в диаметре) — покровные клетки, которые делятся на три категории: экзопинакоциты, формирующие экзопинакодерму (наружный слой губки); базопинакоциты, формирующие базопинакодерму (основание губки, где она крепится к субстрату) и эндопинакоциты, формирующие эндопинакодерму (внутренний слой губки).
Мезенхимальные клетки (18 мкм в диаметре) заполняют внутреннее пространство (мезохил) между камерами хоаноцитов и пинакоцитами. К мезенхимальным клеткам относятся стволовые и скелетогенные клетки губок.
Как видно, перечень клеток у губок невелик, но они обладают генами, кодирующими консервативный молекулярный механизм, обнаруженный в нейронах, мышцах и в специализированных типах клеток билатеральных животных*.
Билатеральные животные* — существа, для которых характерно наличие передне-задней оси тела, системы органов и четко определенной или истинной эндодермальной мезодермы (средний зародышевый листок у многоклеточных животных), которой нет у губок и кишечнополостных.Еще одним любопытным фактом про губки является то, что они демонстрируют различные формы медленных движений, включая ритмические сокращения, которые промывают систему каналов и выводят мусор. Однако клетки с интегративными сигнальными функциями у губок неизвестны, а генетическая подпись и взаимоотношения большинства типов клеток неполны. Следовательно, губки могут быть отличным кандидатом для изучения ранней эволюции определенных типов клеток, в том числе и нейронов.
Результаты исследования
С помощью секвенирования РНК ученые исследовали генетически различимые типы клеток у губки вида Spongilla lacustris (озерная бадяга).

Внешний вид губки Spongilla lacustris.
мРНК было выделены и идентифицированы из отдельных клеток диссоциированных 8-дневных молодых особей S. lacustris. На этой стадии развития у них имеются все морфологические особенности зрелой особи, в том числе многочисленные камеры хоаноцитов, хорошо развитая система каналов и дифференцированный мезохил.
В результате секвенирования были получены данные 10106 отдельных клеток S. lacustris, экспрессирующих 26157 генов из 39552 в транскриптоме S. lacustris.
Используя кластеризацию графов (алгоритм Лувена), было выделено 42 кластера клеток с различными сигнатурами экспрессии генов. Проецирование этих данных в двумерное пространство показало наличие центрального кластера, которого исходили все оставшиеся (1D).
Гены-маркеры для этого кластера включали Noggin, Musashi1 и Piwi-like, которые разграничивают плюрипотентные стволовые клеткообразные археооциты*.
Археооциты* — амебоидные клетки, обнаруженные в губках. Они тотипотентны и выполняют различные функции в зависимости от вида. Структура этих клеток соответствует структуре стволовых клеток, поскольку они содержат большое количество цитоплазмы, что помогает клеткам изменяться в соответствии с их функцией.Экспрессия маркеров археоцитов постепенно снижалась с расстоянием от центрального кластера до периферических кластеров (1E). А кластеры, соединяющие центр и периферию, демонстрировали профили экспрессии, промежуточные между профилями экспрессии археоцитов и периферических кластеров (1F), с несколькими уникальными маркеры.
Но вот кластеры на периферии демонстрировали гораздо более уникальные профили экспрессии, состоящие как из факторов транскрипции, так и из эффекторных генов. Это указывает на присутствие различных программ дифференцировки для этих клеток.
Основываясь на этих наблюдениях, ученые предположили, что «ветви», исходящие из центра, являются своего рода траекториями, идущими от стволовых клеток археоцитов через коммитированные предшественники к дифференцированным клеткам на периферии. Соответственно, периферические кластеры представляют собой дифференцированные типы клеток губки.
В четырех случаях конечные точки траектории включали пару тесно связанных кластеров с сотнями генов, специфичных для этой пары, но и с относительно небольшим количеством генов, специфичных для каждого отдельного кластера. Из этого следует, что эти пары представляют собой различные состояния одного и того же типа клеток.
Затем были выбраны гены-маркеры для каждого кластера и проведена визуализация их экспрессии с помощью одномолекулярной флуоресцентной гибридизации in situ (smFISH от single-molecule fluorescence in situ hybridization). Этот метод позволил получить пространственную карту стволовых клеток, предшественников развития и дифференцированных типов клеток во всем организме.
in situ* — рассмотрение явления именно в том месте, где оно происходит.Зонды, направленные против маркеров археоцитов, пометили популяцию крупных мезенхимальных клеток с выступающим ядрышком (1G). Периферические кластеры 10 и 11 были идентифицированы как хоаноциты и апопиларные клетки на основании экспрессии актин-связывающего белка виллина (1G). Кластеры 8 и 9, которые занимают промежуточное положение между археоцитами и хоаноцитами, показали повышенную регуляцию многих маркеров пролиферации, включая Pcna. Эти кластеры были идентифицированы как хоанобласты.
Методика, описанная выше, позволили отнести все периферические кластеры к 18 типам дифференцированных клеток. По словам ученых, это дало полную классификацию популяций клеток губки.
Основываясь на данных из предыдущих исследований, ученым удалось извлечь пинакоциты и несколько ранее описанных типов мезенхимальных клеток, таких как склероциты, которые производят кремнистые спикулы, и клетки, которые транспортируют спикулы. Однако были и клетки, о которых ранее никто не писал, т.е. они были обнаружены в ходе этого исследования впервые.
Ученые отмечают, что типы клеток у большинства животных организованы в отдельные семейства, которые разделяют экспрессию регуляторных и эффекторных генов. Их идентификация стала возможной благодаря относительно новому методу иерархической кластеризации.
Чтобы изучить взаимосвязи типов клеток у губок, сначала был использован анализ взвешенных сетей коэкспрессии генов (WGCNA от weighted gene coexpression network analysis), позволяющий идентифицировать наборы генов, которые одинаково коэкспрессируют в разных типах дифференцированных клеток. С помощью WGCNA было выявлено 28 различных наборов генов, некоторые из которых указывали на уникальные типы клеток, тогда как другие представляли группы типов клеток. Это значит, что имеет место иерархическая организация.
Дабы удостовериться в этой гипотезе, ученые применили статистическую модель, которая оценивала, имеют ли взаимосвязи между типами клеток, основанные на экспрессии генов, иерархическую организацию или же это всего лишь случайность в пределах погрешности. Этот метод присваивает оценку «чистоты» для каждой комбинации четырех типов клеток (тетрада) и оценивает p-значение с использованием рандомизированных наборов данных. В результате было обнаружено, что в паттернах экспрессии генов среди типов клеток действительно преобладает иерархическая структура. При этом 60% тетрад типов клеток демонстрируют более высокую иерархию, чем если бы это было случайностью. Другими словами, иерархическая система присутствует намеренно.
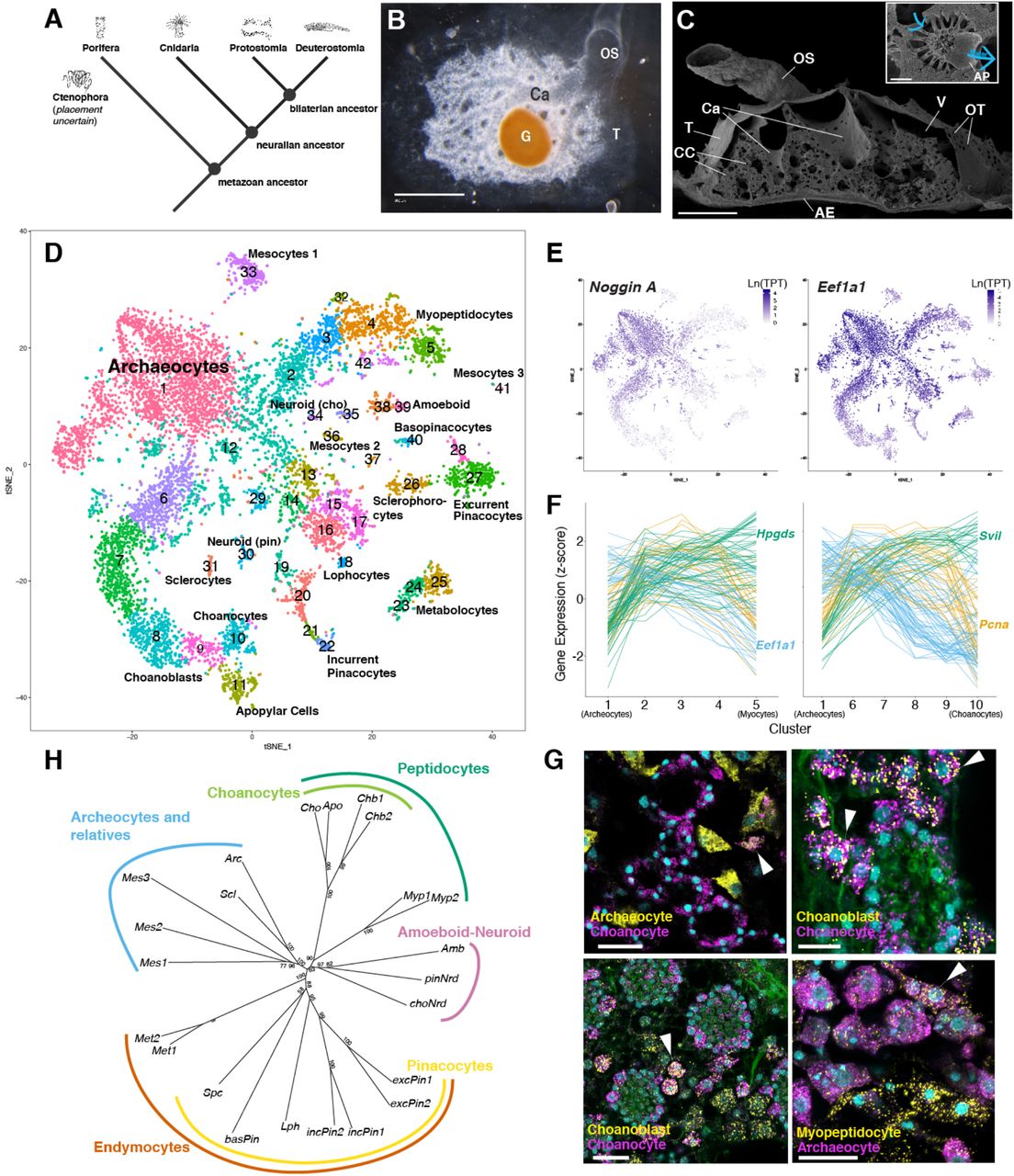
Вышеописанные данные позволили создать дерево типов клеток (1H), демонстрирующее четыре хорошо определяемые клады дифференцированных типов клеток. К ним относятся расширенные семейства пинакоцитов и хоаноцитов, а также два семейства преимущественно мезенхимальных клеток, одно из которых содержит иммунные и нейроидные клетки, а другое — склероциты и неизученные типы клеток мезоцитов.
Изображение №2
Семейство типов клеток, связанных с пинакоцитами, ученые назвали «эндимоцитами» (от греческого ἔνδυμα — подкладка, одежда), которые фомируют внешний слой губки и форму ее тела.
Среди эндимоцитов входящие в состав пинакоциты составляют оба наружных слоя, внутренний слой губки и слой оскулюма* (2A-2D).
Оскулюм* — отверстие на теле губки, находящееся обычно на противоположном от подошвы конце, является частью фильтрационной системы.Экскурсионные пинакоциты выстилают экскурсовые каналы и внутренний слой оскулюма (2E), формируя всю экскурсовую систему. Наконец, базопиноциты составляют базальный эпителиальный слой, прикрепленный к субстрату (2G).
Мезенхимные члены семейства эндимоцитов, включая лофоциты, секретирующие коллаген (2H), склерофороциты (2I) и метаболоциты (2J), чаще всего наблюдались около базального слоя и на периферии губки, часто вблизи пинакоцитов.
Анализ генной онтологии модулей (кластеров коэкспрессирующихся генов) генов эндимоцитов обнаружил сильное обогащение для передачи сигналов Wnt и TGF-β, а также модуль сократимости белка мышечных волокон актомиозина (2K). Также наблюдалась экспрессия Srf (сывороточный фактор ответа), который является главным регулятором сократительного аппарата. В добавок обнаружилась экспрессия цистеина и богатого глицином белка (Csrp1/2/3), кодирующих кофакторы Srf. Из этого следует, что губки могут содержать консервативный регуляторный модуль для сокращения актомиозина (2K).
В предыдущих исследованиях было показано, что сигнальные молекулы, такие как ГАМК, глутамат и оксид азота, изменяют ритмические сокращения. Подтверждением этому стало обнаружение паралогов* метаботропного рецептора ГАМК-B, специфически используемых в отдельных эндимоцитах.
Паралог* — один из двух генов, возникших в результате дупликации исходного гена.Пинакоциты также специфически коэкспрессируют синтазу оксида азота, которая катализирует синтез NO, а также Gucy1B2 — рецептор NO (2K). NO вызывает расслабление эндотелиальных мышц позвоночных и участвует в регуляции сокращений губок.
Затем ученые решили охарактеризовать сокращения у S. lacustris и роль NO в регуляции этого поведения. Сначала была создана общая модель сокращений S. lacustris. Индивидуальный цикл сокращения у S. lacustris состоит из семи различных стадий, каждая из которых управляется скоординированными морфологическими изменениями во входящей (поступление воды), исходящей (выброс воды) и оскулярной тканевых системах.
Видео №1: сокращения S. lacustris.
Во время первых трех стадий сокращение оскулята и входной системы заставляет воду наполнять каналы. На стадиях 4 и 5 система экскурсов сжимается, вытесняя воду через оскулюм. На заключительной стадии все системы возвращаются в состояние покоя.
Затем была проверена реакция молодняка Spongilla на NO. Для этого ювенильных губок обработали NOC-12 (низкомолекулярный донор NO) и ODQ (ингибитор NO-рецептора Gucy1b2) (2L-2M).
Добавление NO приводило к немедленному сильному сокращению входящей системы (2L), наряду с одновременным расширением системы экскурсовых каналов, подобно тому, что наблюдается на начальных стадиях эндогенного сокращения.
Видео №2: применение NOC-12 на губках S. lacustris.
В постоянном присутствии оксида азота текущая система оставалась в сжатом состоянии, претерпевая серию коротких импульсных сокращений. Полная релаксация произошла только после удаления оксида азота промыванием, что также привело к возврату систем в состояние покоя (2L).
Эффект от ODQ был менее заметныv, хоть и наблюдался эффект расслабление систем эксклюзионных каналов (2M), который утихал только после промывания (т.е. удаления ODQ).
Экспрессия указывает не только на функцию сокращения у эндимоцитов, но и на то, что они участвуют в ощущениях, формировании скелета, метаболизме и защите.
Видео №3: применение ODQ на губках S. lacustris.
При этом пинакоциты экспрессируют одиночные ортологи Six и Pax — основных гомеодоменные факторы консервативной Six/Pax регуляторной сети. Также пинакоциты экспрессируют датчик синего света Cryptochrome. Следовательно, эти клетки могут быть светочувствительны.
Самым отличительным членом семейства эндимоцитов являются недавно обнаруженные метаболоциты — большие мультиполярные мезенхимальные клетки, которые могут контактировать с пинакоцитами. Метаболоциты специфически активируют полный набор ферментов, опосредующих гликолиз и пентозофосфатный путь, и экспрессируют гликогенфосфорилазу и пируватдегидрогеназу. Это указывает на то, что они являются ключевыми регуляторами энергетического метаболизма в губке. Кроме того метаболоциты экспрессируют гены nox, кодирующие субъединицы НАДФН-оксидазы, которая продуцирует активные формы кислорода для борьбы с бактериями во время фагоцитоза.
Изображение №3
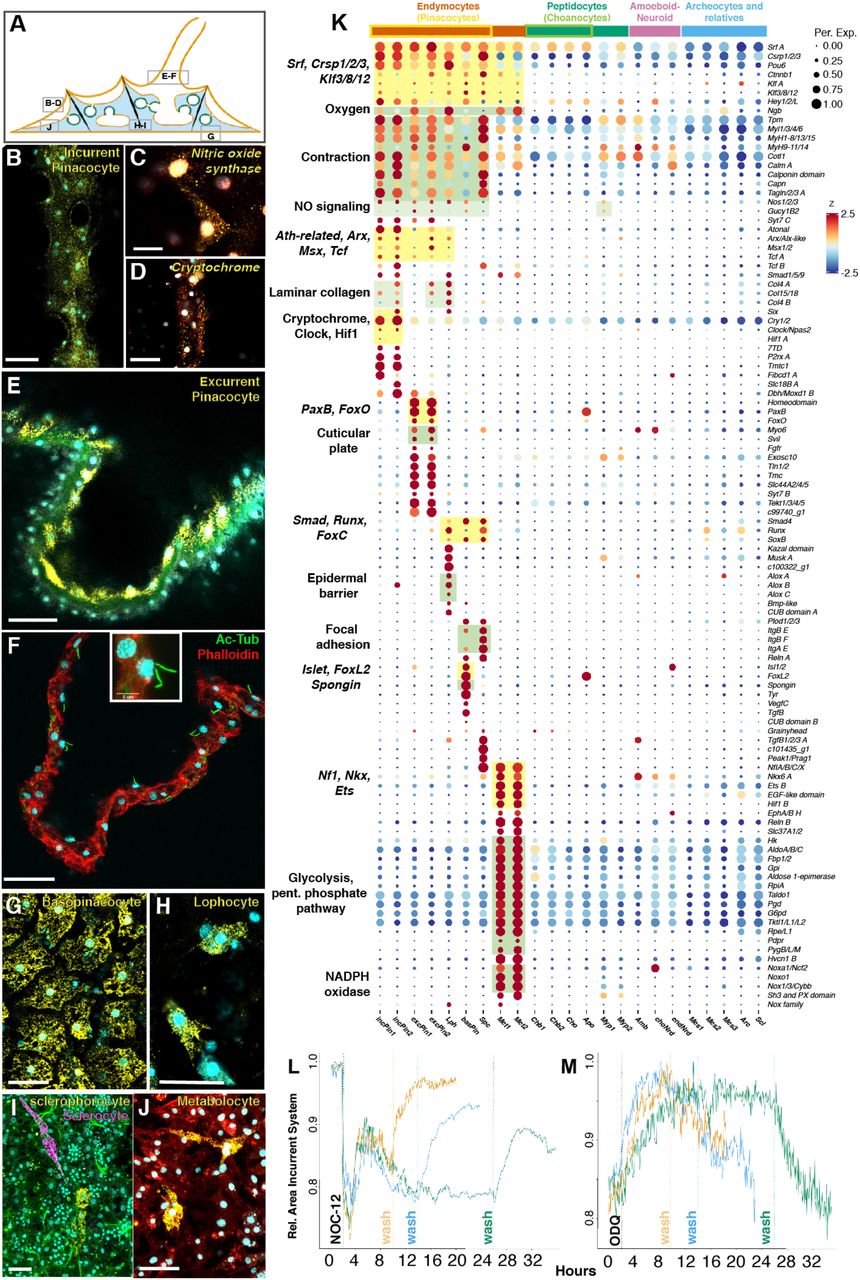
Пептидоциты — семейство клеток, связанных с хоаноцитами (от греческого Πέπτειν — переваривать). Они являются пищеварительными клетками и экспрессируют рецептивные «постсинаптические» гены.
Пептидоциты включают собственно хоаноциты и близкородственный тип клеток, который с помощью smFISH был идентифицирован как апопиларовые клетки (Apo; 3A-3C). Последние образуют выходную пору камер хоаноцитов (3A) и регулируют потоки воды внутри губки посредством контроля диаметра отверстия.
Также были обнаружены миопептидоциты — многочисленный, но ранее не описанный компонент мезогила (3D-3E). Они образуют длинные выступы, которые контактируют друг с другом и с другими клетками, включая хоаноциты. Среди пептидоцитов они являются уникальными, так как экспрессируют множество сократительных генов (3F), которые регулируют целостность миофибрилл.
Недавние исследования показали, что внутренний объем хоаноцитов в основном состоит из пищевых вакуолей, что делает пищеварение важнейшей функцией. Подтверждением этому было обнаружение того, что все пептидоциты сильно обогащены наборами лизосомных генов, включая лизосомные маркеры биогенеза и пищеварительные ферменты, такие как катепсины. Хоаноциты также экспрессируют факторы транскрипции Rbpj, Klf5 и NK homeobox 6, которые регулируют пищеварительные типы клеток, энтодермальное развитие и метаболизм у других животных.
Также наблюдалась экспрессия шести членов семейства Sorting nexin с Phox и BAR доменами (SNX-PX-BAR), участвующими в макропиноцитозе — нерегулируемой форме поглощения. Это согласуется с тем фактом, что хоаноциты поглощают непищевые частицы (например, латексные шарики) и переносят их в лизосомы.
Изображение №4
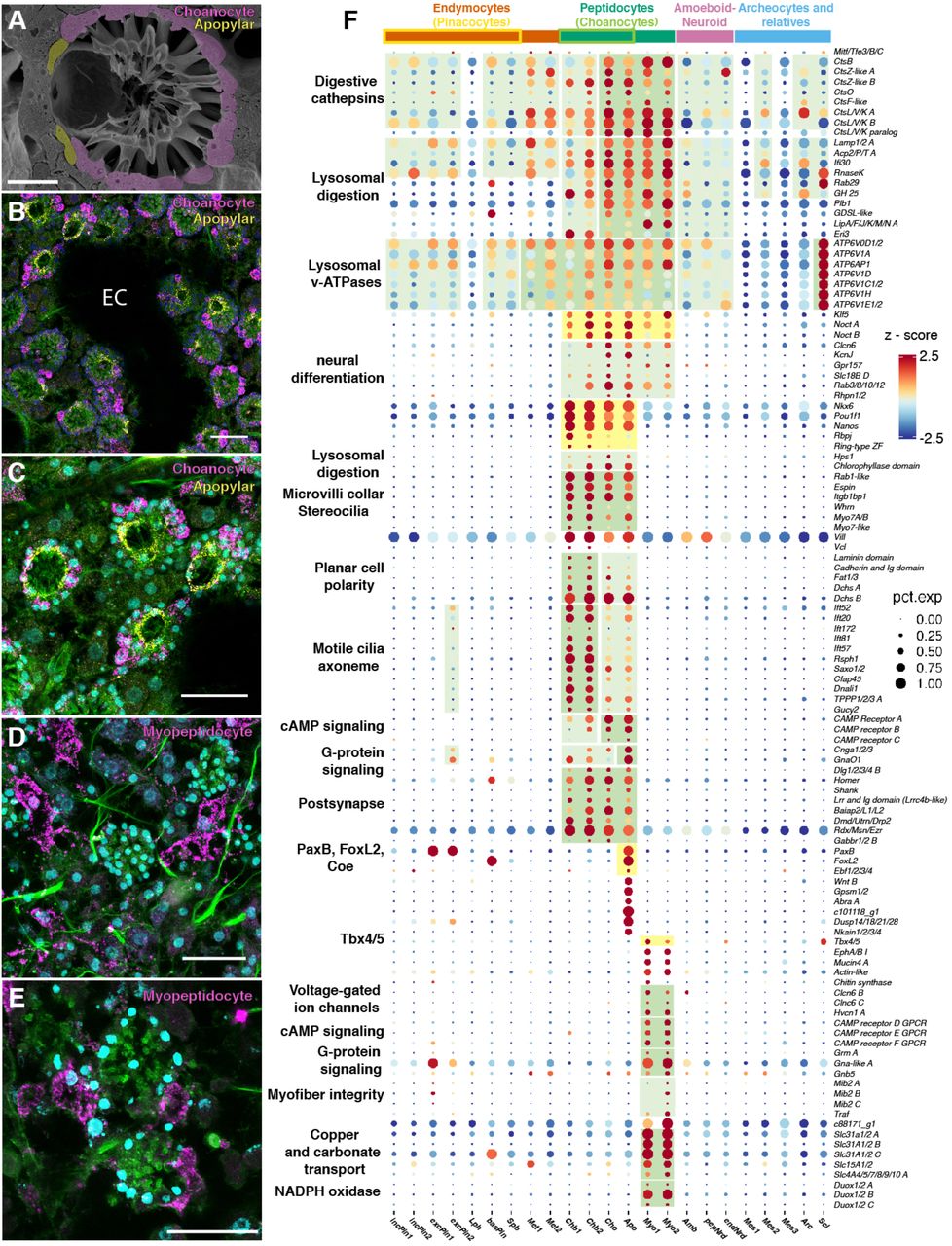
Следующим изученным семейством клеток стало амебоидные нейроиды, включающее в себя три типа мелких клеток, диспергированных в мезогиле. У них нет ядрышка, и они имеют треугольную или многоугольную форму с длинными, подобными филоподиям расширениями (4A-4F). В то время как хоано-нейроидные клетки связаны с камерами хоаноцитов (4A-4D), амебоидные и пинако-нейроидные клетки рассредоточены в мезохиле (4C-4F). Также было выявлено наличие пинако-нейроидных клеток рядом с пинакоцитами (4E).
Факторы транскрипции, специфические для данного семейства, предполагают роль во врожденном иммунитете (4G). Кроме того, амебоциты экспрессируют единственные ортологи губки (Nfat), которые играют важную роль как в адаптивном, так и во врожденном иммунитете.
Видео №4: FIB-SEM данные, показывающие наличие нейроидных клеток в камерах хоаноцитов.
Помимо врожденного иммунитета, наблюдался уникальный «пресинаптический» модуль экспрессии для хоано-нейроидных клеток, что указывает на специфическое взаимодействие и связь с «постсинаптическими» хоаноцитами. Помимо этого, хоано-нейроидные клетки были обогащены генами, участвующими в нейросекреторном переносе и секреции плотных сердцевинных везикул (4G). Из вышеописанных наблюдений можно сделать вывод, что между хоано-нейроидами и хоаноцитами / апопиларными клетками существует взаимодействие.
Дабы проверить эту теорию, ученые проверили тест с контактом между хоано-нейроидами, хоаноцитами и апопиларными клетками. smFISH и снимки конфокальной микроскопии показали, что почти все хоано-нейроидные клетки были связаны либо с камерами хоаноцитов, либо присуствовали в выстилающем слое хоаноцитов, либо располагались в середине камер (4A-4D). Подобная картина напоминает «центральные клетки» губки, которые заселяют камеру хоаноцитов у некоторых обыкновенных губок (Demospongiae), но не у S. lacustris. Они контактируют с окружающими хоаноцитами длинными клеточными выступами. На удивление у S. lacustris все же были обнаружены длинные клеточные отростки выступающие из хоано-нейроидных клеток, выстилающих камеры, а также из более центральных нейроидных клеток, с клеточными выступами, контактирующими с отдельными хоаноцитами.
Видео №5: FIB-SEM данные, показывающие наличие межклеточного взаимодействия между нейроидными клетками и отдельными хоаноцитами.
Затем был использован сфокусированный ионный пучок, дабы найти хоано-нейроидные клетки в объеме 30 мкм3. Используя машинное обучение, ученые сегментировали весь объем и реконструировали пространственные отношения и взаимодействия всех клеток в камере хоаноцитов, идентифицировав две вероятные хоано-нейроидные клетки, расположенные внутри камеры (5A-5F).
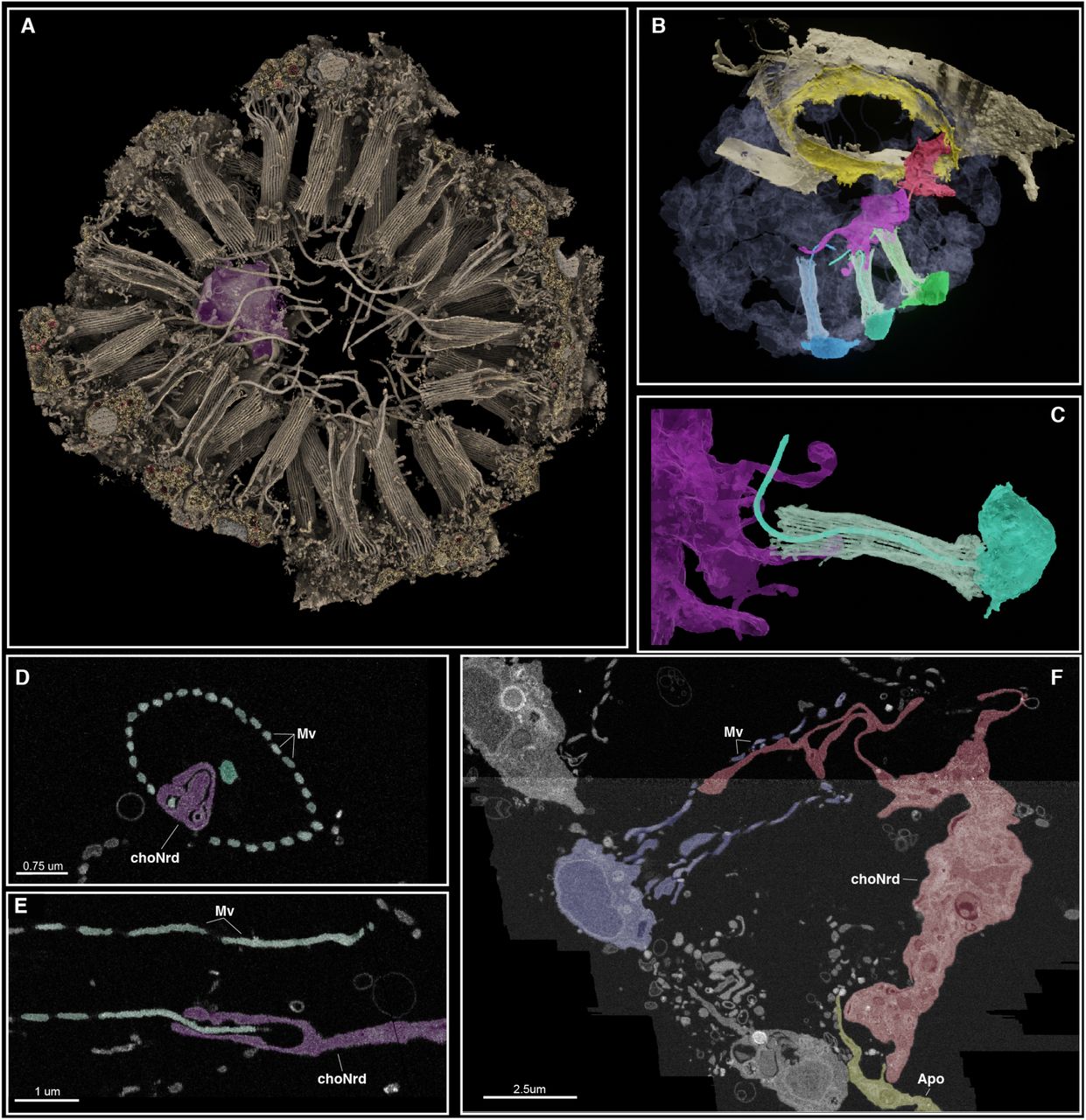
Изображение №5
Первая хоано-нейроидная клетка располагалась в центре камеры (фиолетовая клетка; 5A), а вторая клетка располагалась рядом с апопилярной порой (красная клетка; 5B).
Данная реконструкция показала, что обе нейроидные клетки образуют множественные отростки, каждый из которых направлен к отдельному хоаноциту (5B-5F). Поразительно, что почти все отростки от хоано-нейроидных клеток контактируют и охватывают одну или несколько микроворсинок (5D-5E), причем каждый отросток достигает своего хоаноцита.
Подобная структура вполне может быть современным примером зачаточной стадии развития мозга, которую пришлось пройти многим живым организмам на Земле миллионы лет тому назад.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В данном труде ученые изучили морских губок, являющихся одними из немногих существ, которые не обзавелись мозгами, несмотря на многомиллионную историю существования на планете.
Стоит отметить, что отдельные нейроны в мозге взаимодействуют друг с другом с помощью синапсов. Подобная связь регулируется рядом определенных генов. У морских губок синапсов, естественно, нет, но вот гены, связанные с синапсами, все же есть. Получается парадокс — мозга нет, но гены, необходимые для его функционирования, есть. Зачем они губкам? Полноценного ответа пока что ученые дать не могут.
Однако благодаря дотошному исследованию им удалось установить, что определенные клетки в пищеварительных камерах губок активируют синаптические гены. Таким образом, даже у примитивных животных, лишенных синапсов, синаптические гены активны в определенных частях его тела.
Пищеварительная камера нужна губке, чтобы отфильтровывать пищу из воды. Параллельно она взаимодействует с окружающей средой и, следовательно, с микробами, ее населяющими. Выяснилось, что определенные нейроидные клетки перемещаются по пищеварительной камере, чтобы избавиться от вредоносных бактерий извне. При этом у этих клеток имеются очень длинные отростки, которыми они соединяются с клетками других типов. Другими словами, имеет место «аналоговая» синоптическая система коммуникации между клетками.
Главный вывод исследования в том, что клетки морской губки, регулирующие питание и контролирующие микробную среду, возможно являются предшественниками того, что мы называем мозгом.
В будущем ученые намерены продолжить работать над этим исследованием, так как остается много вопросов, требующих ответов. Несмотря на всю важность мозга и для людей, и для миллионов других существ, нам мало что известно о его происхождении. Однако примитивные организмы, вроде губок, позволяют совершить путешествие в прошлое и понять, как и откуда взялся столь важный орган.
Пятничный офф-топ:
Говоря о морских обитателях и о мозгах, нельзя не упомянуть одно из самых умных существ — осьминога.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?

drWhy
Интеллект в кармане не спрячешь, но он и не обязан ютиться в мозгу — плесень, не имея мозга, сознательно выбирает где ей расти.
Симпатяга с КДПВ мог бы быть одним из братьев Виндоуса, Глинко и Вилли Баранкина из Неверхуда, вылупившимся в океане.
Спасибо за позитивное исследование.
belch84
Не могу удержаться от того, чтобы продемонстрировать устройство мозга одной известной губки
ksr123
Прям сознательно? Или просто благодаря химии?
drWhy
Говорят, что сознательно.