

В ходе эволюции многие представители флоры и фауны приобрели ряд особенностей, которые поражают своей сложностью и приспособленностью к тем или иным условиям обитания. Порой же природа решает применить принцип бритвы Оккама, т. е. пойти по самому простому, а потому и самому очевидному пути. Ярким примером тому являются сосновые шишки, внутри которых имеются семена: если влажность воздуха слишком высока, то шишка как бы закрывается, а если влажность низкая (что хорошо для распространения семян) — шишка раскрывается, словно бутон цветка. Такой простой механизм позволяет соснам минимизировать потерю семян в случае неподходящих для их распространения погодных условий. При этом возникает вопрос — что именно происходит с шишкой во время ее закрытия или открытия? Ученые из Фрайбургского университета (Германия) решили раскрыть эту тайну и провели ряд опытов и наблюдений. Что приводит шишку в движение, как этот процесс протекает, и чем полезны полученные в ходе наблюдений сведения? Ответы на эти вопросы мы узнаем из доклада ученых. Поехали.
Основа исследования
Шишки, являющиеся видоизмененным побегом, присущи многим голосемянным растениям, которые появились на планете порядка 370 миллионов лет тому назад. На данный момент насчитывается более 1000 видов, у каждого из которых шишки обладают определенными особенностями. Любопытно, что секвойи вечнозеленые считаются одними из самых высоких видов деревьев, достигая более 100 м в высоту и более 10 м в диаметре. При этом шишка такого гиганта всего лишь 15-32 мм. А вот у сосны Ламберта (до 70 м в высоту и до 2 м в диаметре) шишки могут достигать 50 см в длину. А вот шишка саговника (до 15 м в высоту) может весить 50 килограмм.

Сравнение шишки секвойи вечнозеленой (сверху) и сосны Ламберта (снизу).
Шишки бывают как мужские, так и женские. В первом случае внутри находится пыльца, после распространения которой шишки опадают. У женских шишек имеется стержень, к которому спирально крепятся чешуйки двух типов (кроющие и семенные). Как понятно из их названий, кроющие чешуйки прикрывают семенные, так как на вторых находятся сами семена. Созревание семян у разных видов занимает от одного до нескольких лет.
Шишки некоторых голосемянных реагируют на влажность воздуха. Для наиболее эффективного распространения семян им нужен сухой воздух, потому при повышенной влажности кроющие чешуйки закрываются, тем самым защищая семенные. Если же влажность подходящая, то шишка раскрывается (1A-1C). Такая методика распространения семян крайне распространена среди деревьев из семейства сосновых (Pinaceae), о которых и пойдет речь в исследовании.
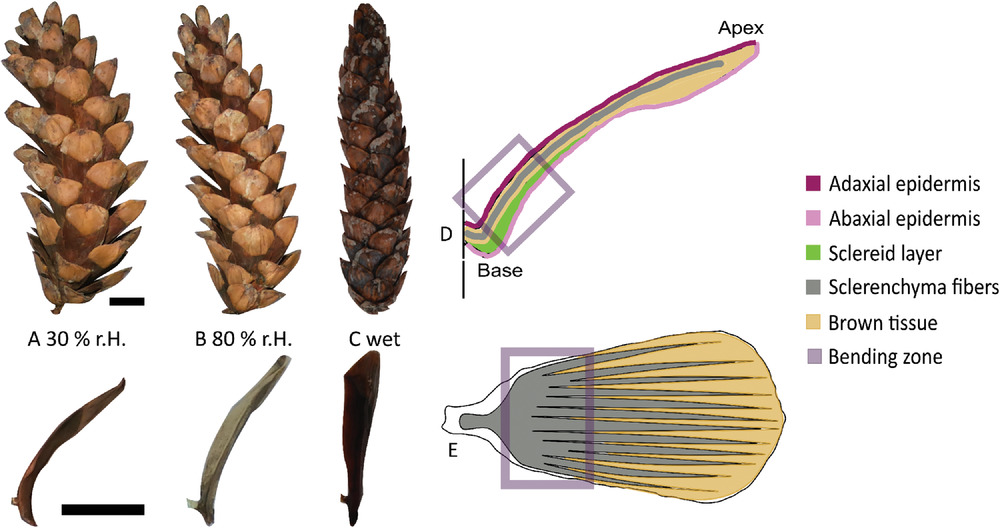
Изображение №1
Переход из «влажного состояния» в «сухое состояние» вызывает пассивное, управляемое водой изгибающее движение индивидуальной чешуйки, что продиктовано различной механикой и свойствами набухания/усадки тканей. Абаксиальный (обращенных к основанию побега) склероидный слой набухает и сжимается в продольном направлении, что обусловлено микрофибриллами целлюлозы, встроенными в клеточные стенки. Предполагается, что слой склеренхиматозных волокон (более адаксиально расположенный, т. е. обращенный к оси побега) является пассивным, резистентным слоем, определяющим, таким образом, деформацию изгиба.
Демонстрация раскрытия чешуек шишки.
Согласно ранее проведенным исследованиям, семенная чешуйка функционирует аналогично биметаллической пластине, но реагирует на влажность, а не на тепло из-за своей функциональной двухслойной архитектуры. Однако эта двухслойная структура присутствует только в самой базальной части чешуи, где активирующий слой (склереиды) и резистентный слой (склеренхиматозные волокна) плотно упакованы и образуют более или менее непрерывные слои.
В других трудах говорится, что подобное объяснение строения является довольно упрощенным в отношении всего масштаба, поскольку оно не включает в себя несколько дополнительных структурных особенностей:
- склеренхиматозные волокна не образуют сплошного тканевого слоя, а отходят в виде тяжей на периферию чешуи;
- нити склеренхимных волокон погружены в матрикс так называемой бурой ткани;
- чешуйки покрыты эпидермисом, который, как самая внешняя ткань, находится в непосредственном контакте с окружающей средой и регулирует поглощение и выделение влаги (1D, 1E).
Механический вклад этих тканей в движение, а также их поведение при абсорбции и десорбции воды еще не были установлены и описаны в научной литературе.
Чтобы произошло движение чешуи, молекулы воды должны диффундировать через эпидермис. Они оседают на поверхности в виде тумана или росы. В то время как туман состоит из мелких капель воды (диаметром около 10 мкм), которые уже находятся в воздухе и могут осаждаться на различных поверхностях, роса образуется только при достижении точки насыщения и конденсации влаги в капли воды. Однако, кроме перенасыщения воздуха или резких перепадов температуры, необходимо учитывать и другие факторы образования росы на поверхностях. Это может включать химические свойства, такие как гидрофильность и/или шероховатость поверхности, как у надкрылий пустынного жука Physasterna cribripes, которые вызывают накопление росы и образование капель воды в пустыне ночью.

Образование капель воды на надкрыльях жука Physasterna cribripes.
Однако, как именно сосновая шишка осуществляет поглощение воды из окружающей среды и, следовательно, распределение внутри чешуек, пока остается загадкой.
Вероятно динамика сосновых шишек обусловлена исключительно изменением влажности и не требует прямого контакта с водой. Диффузия молекул воды обычно зависит от температуры, времени и свойств материала, таких как гидрофильность или пористость.
В недавнем труде «Hydration-induced reversible deformation of the pine cone» ученые исследовали ткани и изгибающее движение чешуек шишек сосны Pinus torreyana (сосна Торрея) с помощью сканирующей электронной микроскопии (СЭМ) и микротомографии (μ-CT). Они обнаружили, что слой склероидов характеризуется градиентом пористости, где высокопористая часть слоя склероидов действует как подушка между нитями волокон склеренхимы и менее пористым слоем склероидов.
В рассматриваемом нами сегодня труде ученые взяли за основу данные своих коллег и объединили их с более подробной информацией о физике и химии поглощения воды, дабы установить ее влияние на движение чешуек.
Результаты исследования
Описание структуры чешуек шишки
Семенная чешуйка сосны гималайской (P. wallichiana) состоит из нитей склеренхимных волокон, склероидных клеток и бурой ткани, инкапсулированных адаксиальным и абаксиальным эпидермисом. Склероидные клетки состоят из удлиненных клеток (2А), в то время как отдельная нить склеренхимы представляет собой плотно упакованный пучок множества мелких волокон (2В).

Изображение №2
Бурая ткань состоит из изодиаметрических клеток (1C, 1D). Адаксиальный эпидермис выглядит довольно гладким (2E), в то время как абаксиальный эпидермис показывает более структурированную поверхность при исследовании с помощью СЭМ (2F).

Изображение №3
Далее ученые реконструировали трехмерную архитектуру целой чешуйки сосны обыкновенной (P. sylvestris) и обнаружили, что чешуйка окружена абаксиальным и адаксиальным эпидермисом (3А). Абаксиальный эпидермис было невозможно отличить от склероидного слоя. Бурая ткань расположена непосредственно под адаксиальным эпидермисом и составляет самую большую часть ткани. Апофиз расположен на вершине чешуйки (3В). Нити склеренхимных волокон погружены в бурую ткань, а потому видны только в случае ее удаления. Нити склеренхимных волокон начинаются как одна большая нить у основания чешуйки, а затем делятся на множество нитей по направлению к вершине (3С).

Изображение №4
Анализ μ-CT позволил сравнить размеры ткани в сухом и влажном состояниях. Угловая разность чешуйки P. sylvestris в сухом и во влажном состоянии составляет ????water = 24° (4A, 4B), а угловая разность чешуйки P. wallichiana при относительной влажности от 80% до 30% и при 23 °С составляет ????80% = 12°.
Вид сверху (4C, 4D) показывает, что нити склеренхимных волокон простираются в виде нитей на периферию чешуйки. Во влажном состоянии слой склероидов был на 27 % толще и на 22 % длиннее в зоне изгиба по сравнению с сухим состоянием. Нити склеренхимных волокон были на 3% тоньше, а на адаксиальной стороне в среднем на 1% длиннее. Это соответствует усадке поверхности адаксиальной чешуи при смачивании (-1% по всей длине и даже -12% в базальной области). Напротив, абаксиальная поверхность при смачивании удлиняется (+14% по всей длине и +18% в базальной области). По объему нити склеренхимных волокон увеличились на 11% при смачивании, а склероид показал увеличение на 16%. Объем бурой ткани при набухании увеличивался на 33%.
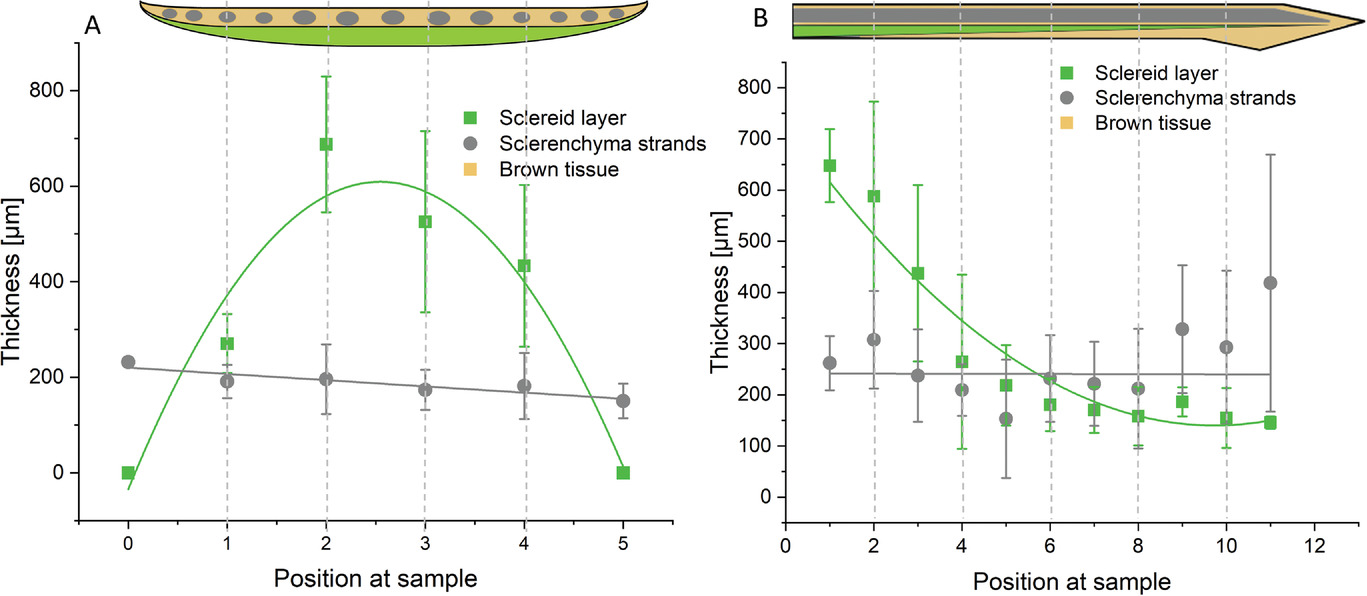
Изображение №5
На 5A и 5B показана поверхность чешуйки в поперечном и продольном сечении. Склероидный слой наиболее толстый в середине и основании чешуи и уменьшается к периферии и вершине. В обоих направлениях толщина нитей склеренхимных волокон остается более или менее постоянной, поскольку в основном более толстые центральные нити склеренхимных волокон остаются дистальными.
Измерения гидратации
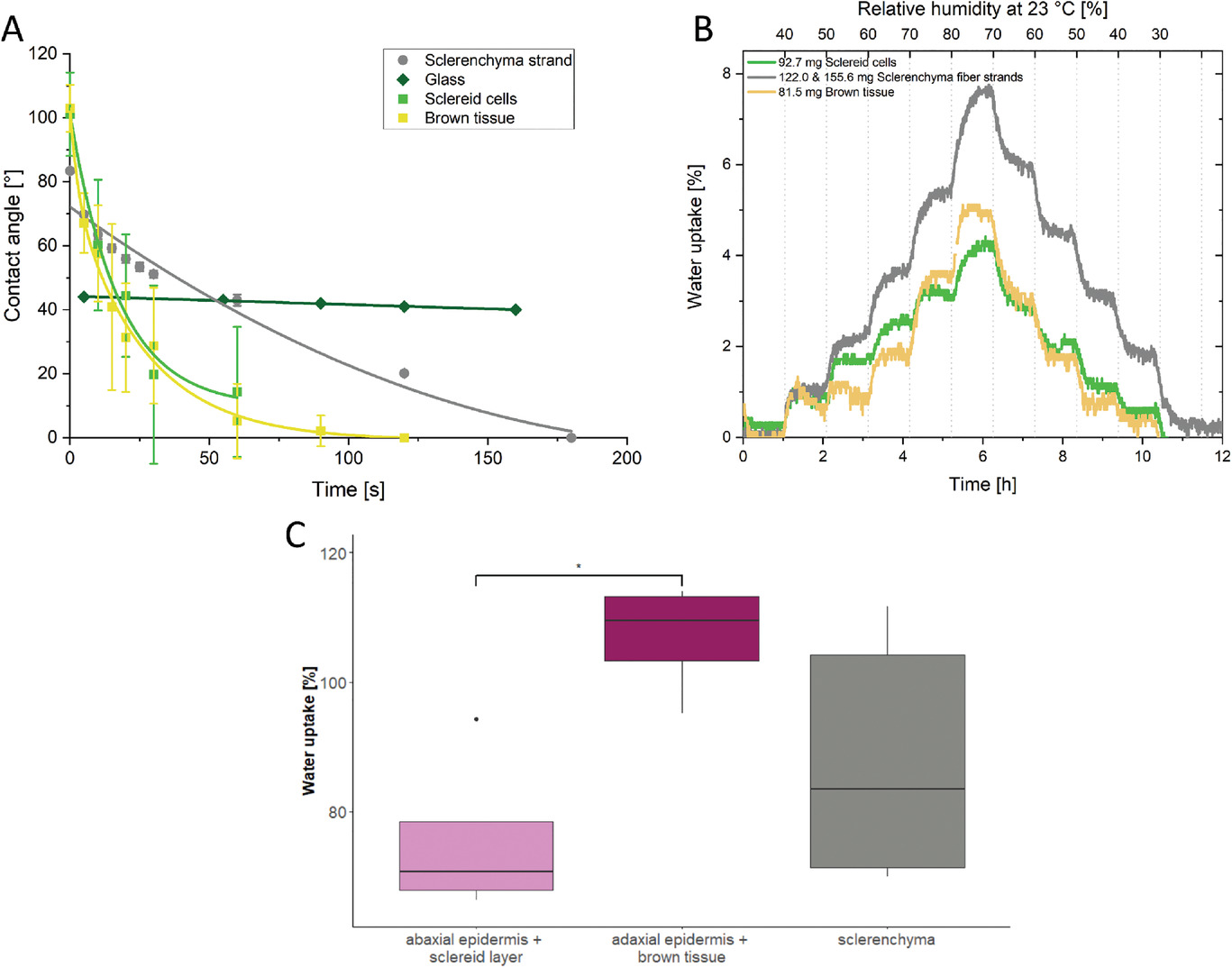
Далее ученые измеряли контактные углы сидячих капель воды в зависимости от времени контакта как на адаксиальном, так и на абаксиальном эпидермисе. Измерения показали, что обе поверхности изначально достаточно сильно гидрофобны (ΘAD, 0 с = 105 ± 6°; ΘAB, 0 с = 119 ± 1°) (розовая и фиолетовая кривая на графиках выше). Примерно через 30 секунд контактный угол уменьшился ниже 90° для обоих образцов. На абаксиальном эпидермисе капля воды объемом 5 мкл полностью растекается в течение 180 секунд почти линейным образом и полностью абсорбируется.
Ход уменьшения контактных углов адаксиального эпидермиса вначале также очень быстрый, но при t = 60 секунд кинетика изменения контактного угла становится похожей на эталонное измерение, где контактный угол изменяется исключительно за счет испарения (зеленая кривая на графике ниже). Даже через 900 секунд капля на адаксиальном эпидермисе показала краевой угол ΘAD, 900 с = 52 ± 18°.

Изображение №6
Выборочно покрывая адаксиальный, абаксиальный или оба эпидермиса интактной чешуи слоем ПММА (оргстекло, PMMA), ученые смогли проанализировать вклад эпидермиса в гидравлическое движение по сравнению с чешуей покрывающего слоя. Угловые изменения 12 чешуек были измерены при увеличении влажности от 30% до 80% и при температуре 23°С. После обработки чешуйки оставляли в покое на 7 часов. Чешуйки без покрытия показали стабильную деформацию изгиба ????NoPMMA = 12.2 ± 4.2°. Достаточно высокое стандартное отклонение связано с широким диапазоном различных размеров чешуек, которые изгибаются в разной степени. Шесть чешуек были покрыты на их адаксиальном эпидермисе, что привело к чуть более низким угловым изменениям ????AD PMMA = 11.5 ± 2.9°. Другие шесть чешуек были покрыты оргстеклом на их абаксиальном эпидермисе, что повлекло за собой уменьшение угловых изменений до ????AB PMMA = 8.5 ± 2.3°.
Далее все оргстеклом покрывали все ранее необработанные стороны всех чешуек. Когда покрывающий слой ПММА был на обеих сторонах чешуйки, водопоглощение значительно снижалось, а угловые изменения падали до ????Compl PMMA = 3.8 ± 2.5°.
Изображение №7
Далее были проведены измерения кинетики развития контактного угла волокон склеренхимы, слоя склероидов и бурой ткани (7А). Чтобы узнать больше о пути воды внутри чешуек, учитывалась не только скорость, но и степень поглощения воды. Для этого были проведены гравиметрические измерения в течение 1 часа (7B).
Краевой угол бурой ткани (7А) начинался при ΘBT, 0 с ≈ 100°. Он экспоненциально уменьшался, и капля поглощалась в течение 120 секунд. Кроме того, бурая ткань (82 мг, 23 °C, 80 % влажности) абсорбировала 5 % массы воды (≈ 4.1 мг; 7B). Бурая ткань достигала равновесия в аспекте поглощения воды примерно через 20 минут.
Контактный угол слоя склероидов (зеленая линия на 7А) также начинался примерно с ΘSC, 0 с ≈ 100° и также демонстрировал экспоненциальное уменьшение, указывающее на абсорбцию, но немного медленнее, чем у бурой ткани. Однако из-за набухания и деформации образца краевой угол можно было измерить только до t = 60 с после нанесения капли (ΘSC, 60 с ≈ 18°).
Склероидные клетки (93 мг, 23 °C, 80% влажности) поглощали 4.5% массы воды (≈ 4.2 мг; 7B). Набухание также достигло равновесия примерно через 20 минут, но время уравновешивания увеличивалось, если влажность была выше.
Контактный угол нитей склеренхимных волокон (серая линия на 7А) был гораздо менее гидрофобным и начинался при ΘSF, 0 с ≈ 83°. Хоть медленное уменьшение краевого угла также наблюдалось, через 3 минуты капля полностью поглощалась тканью. Это более медленное поглощение воды также отражается в результатах гравиметрического измерения нитей склеренхимы (серая линия на 7В). В этих измерениях они достигают равновесия медленнее, чем склероиды и бурая ткань. Нити склеренхимных волокон (≈ 140 мг, 23 °C, 80% влажности) поглощали ≈ 7.8% массы воды (≈ 11 мг), что является большей долей среди всех тканей в чешуйке, особенно в диапазоне более высокой влажности. По этой причине равновесное состояние нитей достигалось только спустя 1 час. Другие же ткани достигали такого же состояния значительно быстрее, но поглощали меньше воды.
В качестве дополнительного теста были проведены измерения поглощения воды тканями, полностью погруженными в воду. В этих условиях абаксиальный эпидермис со склероидным слоем увеличился на 71 ± 10.4 мас.% (в среднем с 246 до 421 мг), а адаксиальный эпидермис в сочетании с бурой тканью увеличил свою массу на 110 ± 9.9 мас.% (в среднем с 143 до 290 мг; 7С). Нити склеренхимных волокон поглощали 83 ± 33 мас.% воды, таким образом увеличивая свою массу в среднем с 141 до 288 мг.
Механические свойства
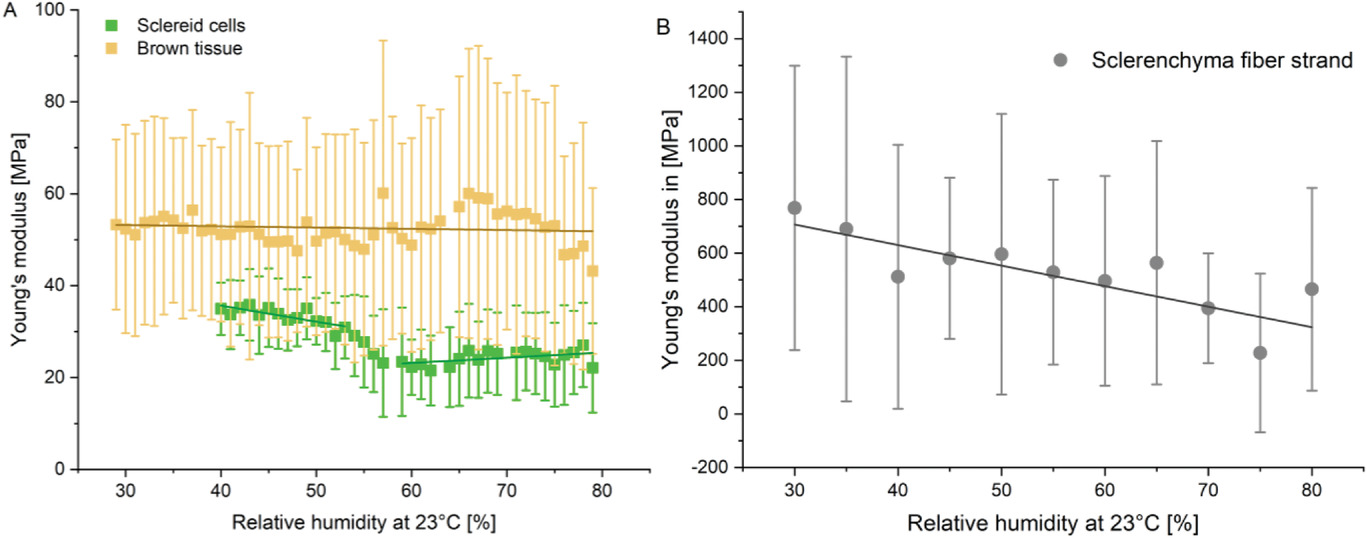
Изображение №8
С помощью атомно-силовой микроскопии (АСМ) были проведены измерения модуля Юнга всех тканей вдоль волокна склеренхимы в зависимости от ориентации нитей и относительной влажности (температура была 23 °C). Как и ожидалось для неоднородных биологических образцов, модули показали сильную дисперсию отдельных значений между 64 точками, измеренными для каждого образца. Однако среднее значение показало четкую тенденцию.
Модуль Юнга склероидного слоя был постоянным при низкой и высокой влажности, но претерпевал почти ступенчатое снижение между 50% и 60% влажности, от ESC, 50% = 35 МПа до ESC, 60% = 25 МПа.
Бурая ткань обладала постоянными механическими свойствами (52 МПа) при влажности от 30% до 70%. Если же влажность превышала 70%, то показатели немного уменьшались до 43 МПа.
Нити склеренхимных волокон, обладающие самым высоким (более чем на порядок выше) модулем Юнга среди тканей шишки, показывали линейное уменьшение показателей при поглощении воды от ESF, 30% = 800 МПа до ESF, 75% = 200 МПа.
Моделирование методом конечных элементов

Изображение №9
На следующем этапе исследования ученые выполнили моделирование методом конечных элементов (изображение №9), что позволило описать модуль Юнга нитей волокон склеренхимы и сравнить их с экспериментально полученными значениями, которые характеризовались высоким стандартным отклонением. Таким образом, можно было смоделировать ситуации, которые не могут быть достигнуты экспериментально (например, сжатие тканей во время сушки от 80% до 30% относительной влажности).
Для моделирования чешуйка была разделена на две зоны. В первой зоне, зоне изгиба (1E), нити склеренхимных волокон расположены очень близко друг к другу, так что между ними почти нет бурой ткани. Эту конфигурацию можно рассматривать как единый однородный слой.
Ученые отмечают, что в отличие от простой модели Тимошенко, в 3D-модели учитывались различная толщина и ширина слоев склероида и склеренхимы (боковые срезы на изображении №9). Во второй зоне ткани не участвуют в сгибательном движении. Таким образом, между концом зоны изгиба и концом чешуйки структура считалась полностью прямой ( все справа от поперечного сечения на 9B). Влажное состояние, то есть согнутое состояние, было установлено в качестве исходной ситуации (9А), а при моделировании отслеживалось высыхание чешуйки (9В).
Моделирование было основано на сокращении слоя склероидов на 22%, наблюдаемом при μ-CT измерении, в то время как образец высушивался от полного погружения в воду до высыхания на воздухе (при моделировании это принималось за объемную усадку), и на измеренных модулях Юнга склероидного слоя. Кроме того, форма чешуйки во влажном состоянии настроена на угловое изменение 0° и сжатие 0%, а в сухом состоянии ????dry = 24.4° для углового изменения и 22% для сжатия.
Результаты моделирования показали, что модуль Юнга склеренхимы должен был составлять 1170 МПа, чтобы достичь углового изменения 24.4°. Предполагая, что сокращение склероида, а также модуль Юнга волокон склеренхимы увеличиваются линейно при сушке (модуль Юнга при полном высыхании равен 70 МПа), удалось построить график развития углового изменения (график ниже).
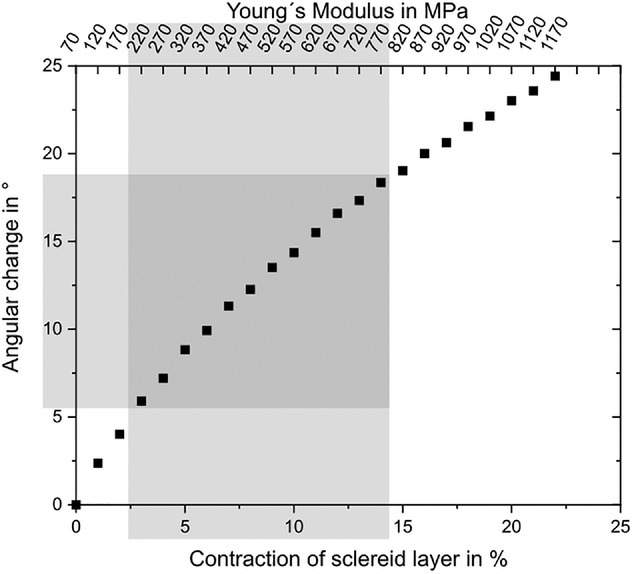
Изображение №10
Для проверки сильного рассеяния измеренных модулей Юнга волокон склеренхимы между 30% относительной влажности (≈180 МПа) и 80% относительной влажности (≈800 МПа) угловое изменение, полученное в результате моделирования (выделено серым цветом на графике выше), сравнивалось с измеренным значением. Как моделирование, так и измерения показали 12° углового изменения.
Таким образом, можно сделать вывод, что при сушке от 80 до 30 % слой склероидов сокращается примерно на 11 %, а модуль Юнга волокон склеренхимы увеличивается на 620 МПа.
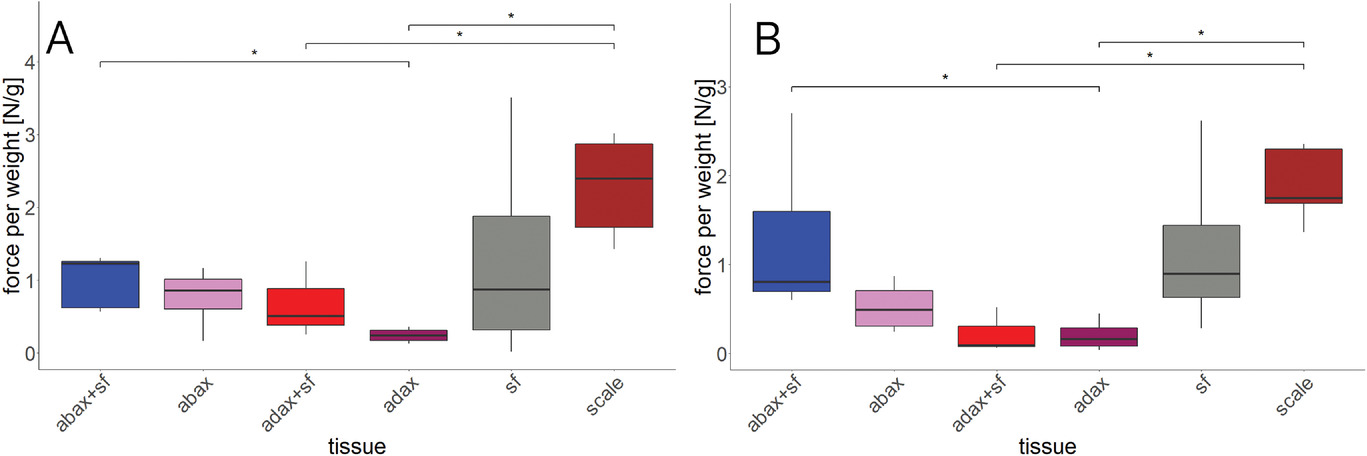
Изображение №11
Измерения силы (графики выше) показали, что необработанные целые чешуйки массой 548 ± 0.06 мг развивали силу около 1.3 Н (2.4 Н/г) при набухании и 0.9 Н (1.8 Н/г) при сушке.
Абаксиальный эпидермис со склероидным слоем и нитями склеренхимных волокон показал 1.2 Н/г при набухании и 0.8 Н/г при высыхании. Такой же образец, но уже без нитей показал 0.9 Н/г при набухании и 0.5 Н/г при сушке.
Образцы адаксиального эпидермиса, где нити склеренхимы еще были прикреплены, показали 0.5 Н/г при набухании и 0.2 Н/г при высыхании. Адаксиальный эпидермис с бурой тканью генерировал 0.2 Н/г при набухании и 0.1 Н/г при высыхании. Нити склеренхимных волокон генерировали 0.9 Н/г как при набухании, так и при высыхании.
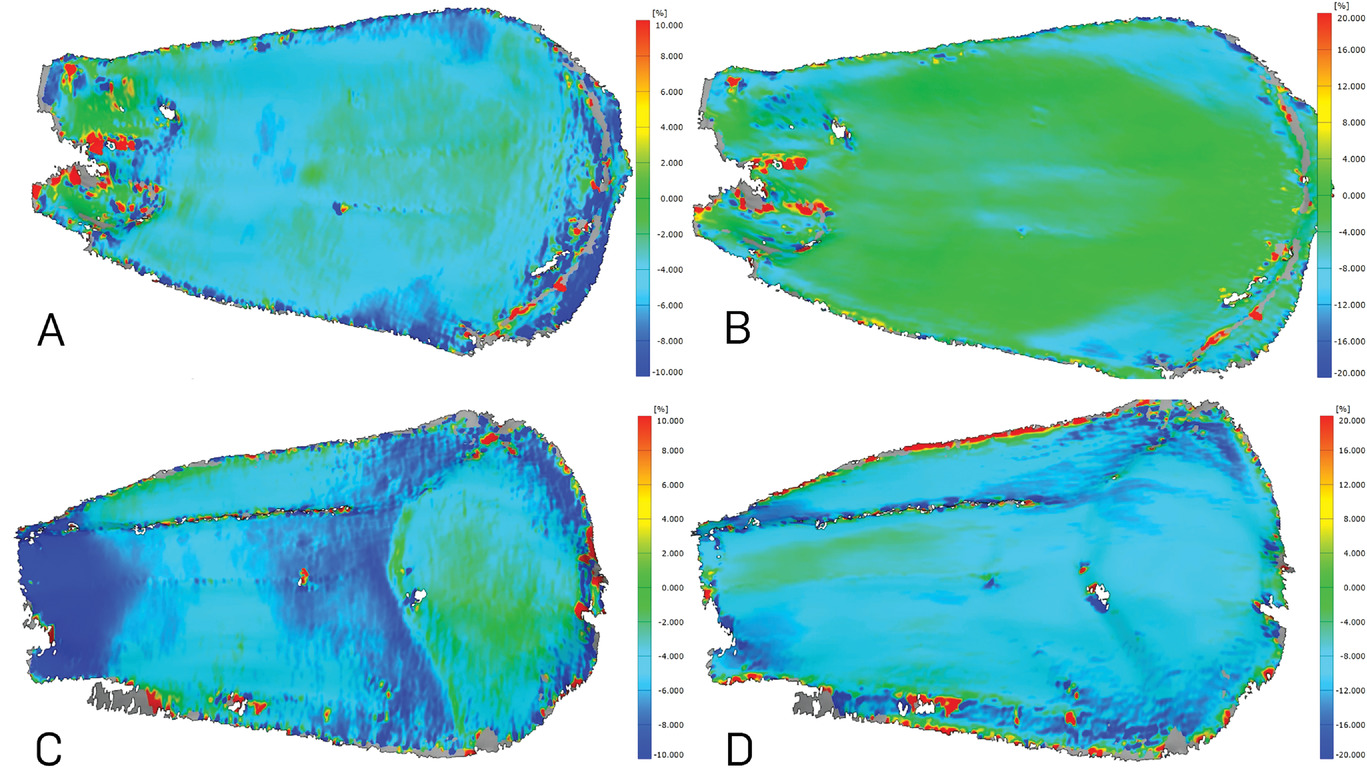
Изображение №12
Используя корреляцию 3D-изображений (3D-DIC), повторяющиеся модели деформации можно было распознать во всех исследованных чешуйках во время движения, вызванного сушкой. Из-за выраженной деформации изгиба вершина чешуйки являлась наиболее смещенной областью. На абаксиальной поверхности возникали в основном отрицательные продольные деформации, которые концентрировались в самой базальной части чешуйки (12А) и уменьшались до -10% и более в процессе движения. На адаксиальной поверхности возникли отрицательные, а также положительные продольные деформации (12С). Особенно в базальной области во время сушки можно было наблюдать несколько пятен с положительной деформацией. Поперечная деформация около -8% наблюдалась на абаксиальной стороне чешуйки (12В). На адаксиальной стороне почти не было поперечных деформаций (12D).
Выводы исследователей
Результаты исследования показали, что чешуйки сосновых шишек изгибаются из-за сложного взаимодействия различных тканей, каждая из которых вносит свой вклад в движение. Изучение кинематики смачивания, гравиметрического водопоглощения и модуля Юнга в зависимости от влажности позволили создать подробную модель поглощения и распределения воды в сосновой шишке. Полученная модель является рассширенным вариантом модели Тимошенко для биметаллических пластин, которая описывает изгибающее движение.
Результаты экспериментов показали, что поглощение воды в основном происходит через абаксиальный эпидермис чешуйки. Поскольку поверхность изначально шероховатая и довольно сильно бороздчатая, контактирующая капля воды находится в состоянии Кэсси-Бакстера с воздушными карманами в канавках под ней. Это приводит к тому, что капля, которая лишь на короткое время соприкасается с чешуйками, скатывается.
При длительном контакте с водой поверхностный слой начинает набухать, краевой угол на шероховатых поверхностях уменьшается, что приводит к переходу от смачивания Кэсси-Бакстера к смачиванию Венцеля. Это позволяет воде растекаться по поверхности чешуек. Затем вода поглощается бурой тканью и нитями склеренхимных волокон, которые также распределяют воду в чешуйке. Нити склеренхимных волокон при этом сильно размягчаются. После поглощения воды склероидным слоем он расширяется в продольном направлении, так как в других направлениях ему препятствует уплотняющий целлюлозный «корсет» в клеточных стенках. Нити склеренхимных волокон позволяют и усиливают изгиб за счет размягчения, а слой склероидов выполняет изгиб за счет расширения.
В отличие от традиционной двухслойной модели, состоящей из набухающего в воде («активного слоя») и набухающего в меньшей степени («пассивного слоя»), чешуйка шишки состоит из нескольких слоев, каждый из которых поглощает воду. При поглощении воды нити склеренхимных волокон сильно размягчаются и, таким образом, «разблокируют» изгибающее движение, которое затем инициируется одномерным набуханием «укрепленных корсетом» клеток в слоях склероидов. Таким образом, коллективное участие всех тканей в одном и том же процессе, т. е. поглощении воды, приводит к раскрытию слоев и изгибанию.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня труде ученые уделили внимание сосновым шишкам. Эти, на первый взгляд, непримечательные объекты оказались довольно интересными с точки зрения механики и, в частности, кинематики.
Сосновые шишки, хранящие в себе семена, раскрываются для их распространения только при подходящих погодных условиях, а именно при низкой влажности. Если же влажность слишком высока, то чешуйки шишки смыкаются, предотвращая проникновения влаги внутрь, что может испортить семена.
Следовательно, ткани чешуек реагируют на влажность, запуская сложный механизм движения самой чешуйки. Любопытно и то, что в этом процессе участвуют и те ткани, которые раньше считались пассивными — нити склеренхимных волокон. Эти нити действительно очень жесткие и «неподвижные» в сухом состоянии, но при смачивании именно они запускают процесс закрытия шишки.
Еще одно важное замечание, сделанное учеными, заключается в том, что хоть все ткани чешуйки и состоят из одних и тех же химически идентичных материалов, их расположение на чешуйке разительным образом отличается. Следовательно, в процессе раскрытия/закрытия более значимую роль играет не химия, а скорее механика чешуек.
По словам ученых, их открытия не только позволяют нам лучше понять, какие химические и физические процессы протекают в природе, но и являются фундаментальными знаниями для создания в будущем более эффективных гигроморфных систем. Кроме того, подобного рода исследования показывают в очередной раз, что природа является отличным источником вдохновения не только для живописцев, но и для ученых.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Комментарии (4)

vviz
09.06.2022 17:10А почему для распростронения семян нужна пониженная влажность? Разве они должны разносится ветром? Разве для всего растительного влажность не один из важнейших факторов выживаемости? Орех, извините, тупо падает из шишки вниз и лежит там, пока его кто-нибудь не съест (кабан) или не прикопает (белка, птица). Какое распространение в зависимости от влажности?
Влажность может испортить семена? Простите, а почему семена не прорастают прямо в шишке? Может потому, что для проростания им нужно оказаться именно во влажной среде?
Объясните мне, плз, популярно.
NEIR0N
похоже что студенту когда то дали какую о тему исследования ради исследования. и так и пошло все, в итоге по всей видимости будет дисер по шишкам))) сомнительная тема для исследования на мой взгляд.
Mimizavr
Вспоминается знаменитое: «Червяк такой длинный, а жизнь такая короткая!»
Wizard_of_light
Ну фиг знает, мне кажется, в каких-нибудь системах климат-контроля такой "шишковый автоматизм" пригодился бы.