
 Когда с нами что-то происходит наш мозг фиксирует это, создавая воспоминания. Изменения, которые при этом происходят с мозгом, принято называть энграммами или следами памяти.
Когда с нами что-то происходит наш мозг фиксирует это, создавая воспоминания. Изменения, которые при этом происходят с мозгом, принято называть энграммами или следами памяти.Вполне естественно, что понимание того, как выглядят следы памяти – основной вопрос изучения мозга. Без этого невозможно построить никакую биологически достоверную модель его работы. Понимание строения памяти непосредственно связано с пониманием того, как мозг кодирует информацию и как он ей оперирует. Все это, пока, — неразгаданная загадка.
Еще большую интригу в загадку памяти вносят исследования по локализации воспоминаний. Еще в первой половине двадцатого века Карл Лэшли поставил очень интересные опыты. Сначала он обучал крыс находить выход в лабиринте, а затем удалял им различные части мозга и снова запускал в тот же лабиринт. Так он пытался найти ту часть мозга, которая отвечает за память о полученном навыке. Но оказалось, что память каждый раз сохранялась, несмотря на временами значительные нарушения моторики. Крысы всегда помнили где искать выход и упорно стремились к нему.

Эти опыты вдохновили Карла Прибрама сформулировать ставшую популярной теорию голографической памяти. В соответствии с ней, аналогично оптической голограмме, каждое конкретное воспоминание не находится в каком-то одном месте коры, а присутствует в каждом ее месте и, соответственно, каждое место коры хранит сразу все воспоминания.
Одно время, очень большие надежды в поисках энграмм связывались с синаптической пластичностью. Способность синапсов менять свою чувствительность давала надежду, что через это можно описать все механизмы памяти. Представление о пластичности синапсов привело к созданию искусственных нейронных сетей. Эти сети показали, как нейрон может обучиться узнавать, что-то общее для набора воспоминаний. Но узнавать общее — это совсем не то же, что хранить отдельные воспоминания.
Если вы не занимайтесь непосредственно нейронауками, то скорее всего у вас сложилось впечатление, что у нейробиологов есть множество теорий относительно памяти, но, видимо, нет уверенности, какая из них верна. И так как, скорее всего, эти теории очень сложны, то о них не особо рассказывают в популярной литературе. Так вот, до сих пор, как ни удивительно это звучит, нет ни одной теории памяти. То есть, есть разные предположения о том, что может быть связано с памятью. Но нет никаких моделей, которые бы хоть как-то объясняли, как выглядят энграммы и как они работают.
При этом накоплены огромные знания о биологии нейронов, о проявлениях памяти, о молекулярных процессах, сопутствующих формированию воспоминаний и тому подобное. Но углубление знаний не упрощает ситуацию, а только усложняет ее. Пока о предмете исследований известно не особо много, удобно фантазировать. Полет фантазии не сильно ограничен рамками знаний. Но по мере того, как становятся известны все новые и новые факты, многие гипотезы отпадают сами собой. Придумать же новые, которые находились бы в согласии с фактами становится все труднее.
Когда в науке наступает такая ситуация – это верный признак того, что где-то в самое начало рассуждений закралась фатальная ошибка. В свое время Аристотель сформулировал законы движения. Он, исходил из того, что видел перед собой. Аристотель сказал, что есть два вида движения: естественное движение и принудительное движение. Естественное движение, по Аристотелю, присуще только небесному веществу и только небесные тела могут двигаться без прикладывания силы. Все остальные «земные» тела для движения требуют приложения силы, иначе любое движение рано или поздно должно прекратиться. Почти две тысячи лет это считалось очевидной истиной, так как все остальные видели перед собой то же самое. Но при этом, почему-то, никому за все эти два тысячелетия не удалось построить ни одной работоспособной теории, которая пошла бы дальше утверждений Аристотеля. И только, когда Галилей и Ньютон указали на досадную ошибку Аристотеля, что, дескать, он забыл про силу трения, оказалось возможным сформулировать известные нам законы механики. Затем, правда, был Эйнштейн, но это уже другая история.
Мне кажется, что таким «принудительным движением», сейчас, в нейронауке выступает «нейрон бабушки». По сути, все основные трудности с построением теории памяти связаны с тем, что оказывается очень трудной задачей увязать конкретный нейрон, если приписывать ему функции детектора какого-либо свойства, и память, которая в силу многих причин, не должна быть жестко привязана к определенному нейрону.
Далее я покажу, как могут выглядеть энграммы для случая, когда нейроны лишаются своих «бабушкиных» наклонностей.
В предыдущих частях был описан клеточный автомат, состоящий из однородных элементов. Когда в любом месте этого автомата создается какой-либо узор активности от этого места расходится фронт волны. В каждом месте этого фронта возникает уникальный специфичный, только для этой волны узор.

Если запомнить какой рисунок создает волна, проходя через какое-либо место, то затем можно в том же месте воспроизвести этот же рисунок и запустить из этого места новую волну. В каждом месте, которое на своем пути пройдет эта новая волна, она будет повторять узор исходной волны.
Если составить словарь, состоящий из конечного числа понятий, то каждому понятию можно сопоставить свою уникальную волну. Тогда, в любом месте автомата по узору проходящей волны можно будет определить, какое понятие распространяет эта волна. И из любого места можно будет запустить волну любого понятия если воспроизвести в этом месте фрагмент узора нужной нам волны.
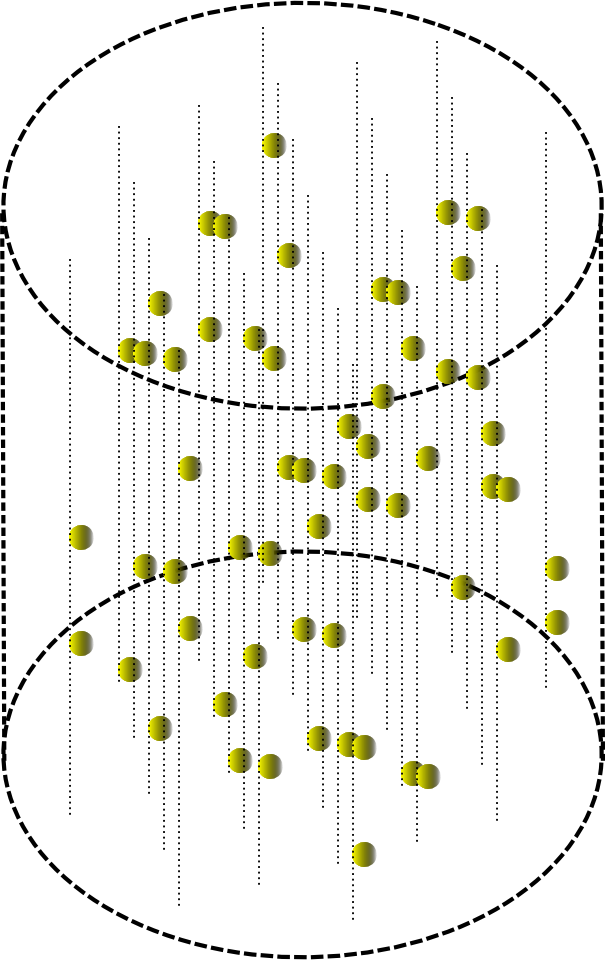
Плоскому авомату можно придать объем.

Прохождение волны в небольшом цилиндрическом объеме тогда будет выглядеть, как показано на рисунке ниже.
Если запустить в автомате информационную волну, а затем запустить волну идентификатора. То можно запомнить картину условной “интерференции” этих волн. Для этого в каждом месте автомата элементами, по которым прошла информационная волна, надо запомнить окружающий их узор волны идентификатора. Такая процедура позволяет запомнить пару «ключ – значение». Если впоследствии запустить в автомат волну идентификатора воспоминания, то элементы автомата воспроизведут узор информационной волны самого воспоминания.
Запомнить информационную пару «ключ – значение» можно, как избирательно в любой малой области автомата, так и глобально во всем пространстве автомата. При глобальном запоминании информация оказывается многократно продублирована по всей площади автомата.
Когда информационное описание состоит не из одного понятия, а из нескольких, то передать такое описание по автомату можно, последовательно распространив информационные волны этих понятий.
В каждом фиксированном объеме автомата прохождение серии волн вызовет смену узоров, каждый из которых можно записать бинарным вектором. Если в описании неважна последовательность понятий, то для одного места автомата бинарные вектора, создаваемые разными волнами можно побитно логически сложить и получить суммарный вектор описания. Этот суммарный вектор при достаточной разрядности сохраняет всю информацию о входящих в него понятиях.
Суммарный вектор имеет высокую разрядность и содержать большое количество единиц. Уменьшить количество единиц и понизить разрядность суммарного вектора можно, рассчитав для него хеш-функцию.
Идентификатор воспоминания, как и само описание, может состоять из нескольких понятий. Тогда и для него можно вычислить соответствующий хеш. При запоминании могут использоваться не исходные коды, а полученные хеши.
Ранее было показано, что для реального мозга возможными кандидатами на роль элементов клеточного автомата могут являться веточки дендритных деревьев нейронов.
Вычисление нейронами хеш-функции дендритных сигналов
Дендритные веточки образуют дендритное дерево (рисунок ниже). Веточки имеют только парные ветвления и не образуют замкнутых циклов.
Модели реальных нейронов (проект EyeWire)
Спайк нейрона возникает, когда деполяризация мембраны на его теле в районе аксонного холмика достигает критического значения. Деполяризация сомы, то есть тела нейрона, происходит, в основном, за счет сигналов от дендритных веточек. Потенциально такими сигналами могут быть токи, возникающие в дендритных веточках и дендритные спайки.
За счет того, что на пути к соме сигналы разных веточек в местах ветвления взаимодействуют между собой, сигналы, доходящие то тела нейрона, оказывается некими функциями сигналов дендритных веточек. Сам мембранный потенциал тела нейрона оказывается функцией сигналов всех веточек его дендритного дерева. Это, кстати, не особо противоречит классическому представлению о формальном нейроне. С той оговоркой, что классический формальный нейрон является простым пороговым сумматором сигналов на синапсах, а мы говорим о достаточно хитрой функции сигналов дендритных веточек.
В описанной трактовке спайк одного нейрона можно смело назвать бинарным результатом хеш-преобразования над сигналами его дендритных веточек. Таким образом, можно говорить о том, что вся картина активности нейронов может быть истолкована, как результат хеш-преобразования активности дендритных сегментах.
Для активации нейрона вся активность, возникающая на дендритном дереве должна уложиться в небольшой временной интервал, составляющий несколько миллисекунд. Если предположить, что за такой интервал и формируется суммарная картина дендритной активности, возникающая после прохождения всех волн сложного описания, то спайки нейронов идеально подходят на роль хеша, связанного с картиной, возникшей на дендритных сегментах.
Для запоминания через интерференцию паттернов необходимо две волны: волна идентификатора и волна значения, то есть сохраняемой информации. В реальной коре эти волны могут распространяться одновременно. При этом, идентификатор сам может быть достаточно сложным описанием. Можно предположить, что в коре хеш для информации и хеш для идентификатора могут формироваться одновременно, но разными нейронами. В принципе, эти нейроны могут быть нейронами разных типов. Самые распространенные нейроны коры – это пирамидальные и звездчатые нейроны. Может так оказаться, что, например, активность пирамидальных нейронов кодирует хеш-функцию для информации, а активность звездчатых хеш-функцию для идентификаторов воспоминаний.
Избранные точки дендрита
Мы пришли к тому, что в каждом месте коры текущая информация может кодироваться сочетанием активности расположенных в этом месте нейронов. Совокупную мгновенную картину их спайков при этом можно воспринимать, как хеш-функцию от соответствующей этим нейронам дендритной активности.
В клеточном автомате для запоминания требовалось, чтобы каждый элемент автомата видел и мог запомнить фрагмент хеш-кода, достаточной длины. Волна идентификатора указывала, какие элементы должны запоминать, а серия информационных волн формировала суммарную картину активности элементов из которой получался тот самый хеш-код, который они должны были запоминать.
В аналогии с мозгом это означает, что каждая дендритная веточка должна видеть активность окружающих ее нейронов и должна иметь возможность ее избирательно запомнить.
Если учесть, что мы хотим, чтобы одна веточка могла запомнить не одну и не две, а тысячи или миллионы различных комбинаций активности нейронов, то задача становится очень интересной.
До сих пор мы, в основном, говорили о дендритных деревьях нейронов, теперь посмотрим на их аксоны. Итак, основной процент нейронов коры приходится на пирамидальные и звездчатые нейроны. Для аксонов этих нейронов характерны сильно ветвящиеся коллатерали. Большая часть синаптических контактов аксона приходится на объем, размеры которого сопоставимы с размером дендритного дерева (рисунок ниже). Такая геометрия аксона обеспечивает то, что сигнал об активности нейрона становится доступен практически всем дендритным веткам этого и других нейронов, находящимся в некоторой окрестности (радиусом порядка 50-70 мкм) этого нейрона.
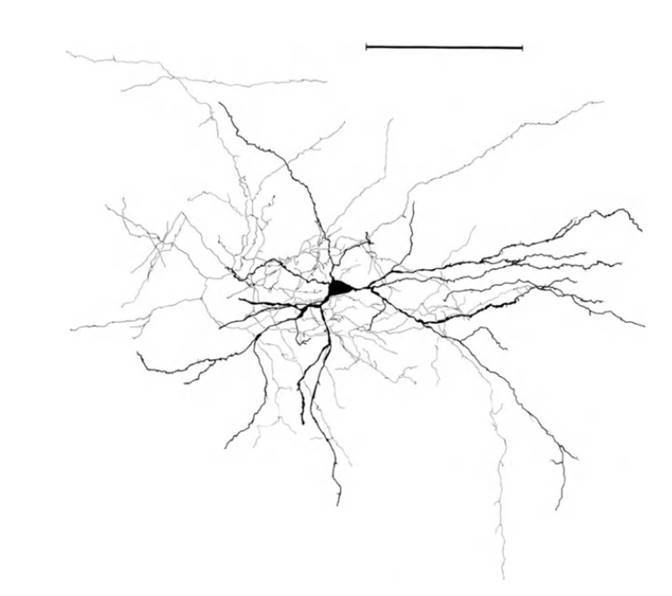
Структура звездчатого нейрона, линейка – 0.1 мм (Braitenberg, 1978)
Доступность сигнала следует понимать в том смысле, что для каждой дендритной ветки поблизости он нейрона будет место где вблизи от нее пройдет аксон этого нейрона. Соответственно, в момент активности нейрона по его аксону распространяется спайк и из всех синапсов, образуемых аксоном, высвобождаются нейромедиаторы. Часть этих нейромедиаторов за счет спиловера, то есть выброса за пределы синапса, может достичь требуемой дендритной ветки.
Вообще, аксоны могут распрстраняться далеко по коре или за ее пределы. Но основное ветвление аксона в большинстве случаев приходится на пространство окружающее сам нейрон. Среднее расстояние между синапсами на дендрите 0.5 мкм. Среднее расстояние между синапсами на аксоне 5 мкм. Число контактов на дендритах равно числу контактов на синапсах. Соответственно, суммарная длина аксона в 10 раз больше суммарной длины дендрита. На ближайшее окружающее нейрон пространство приходится около 6000 его синапсов. Это соответствует длине аксона в 3 сантиметра. Теперь, представьте, что эти 3 сантиметра уложены в сферу радиусом менее одной десятой миллиметра и вы получите представление о характере ветвления аксона. Рядом с любым сегментом дендрита проход множество аксонов соседних нейронов, причем некоторые приближаются к нему не по одному разу.
Синапсы непосредственно окружающие дендритную ветку, как собственные, так и просто, находящиеся рядом, являются источниками внесинаптических нейромедиаторов для этой ветки. Изобразим расположение этих источников вдоль условного сегмента дендрита (рисунок ниже). Для этого совместим источники с дендритом, приблизительно соблюдая их положение по длине дендрита. Занумеруем окружающие дендрит нейроны. Тогда для каждого источника на этой ветке можно указать номер нейрона из окружающего пространства, управляющего этим источником. Каждый из нейронов окружения может иметь сразу несколько контролируемых им источников, случайным образом распределенных по дендриту. Обозначим соотнесение нейронов и источников на дендритной ветке вектором D с элементами di, где каждый элемент – номер нейрона, управляющего источником.
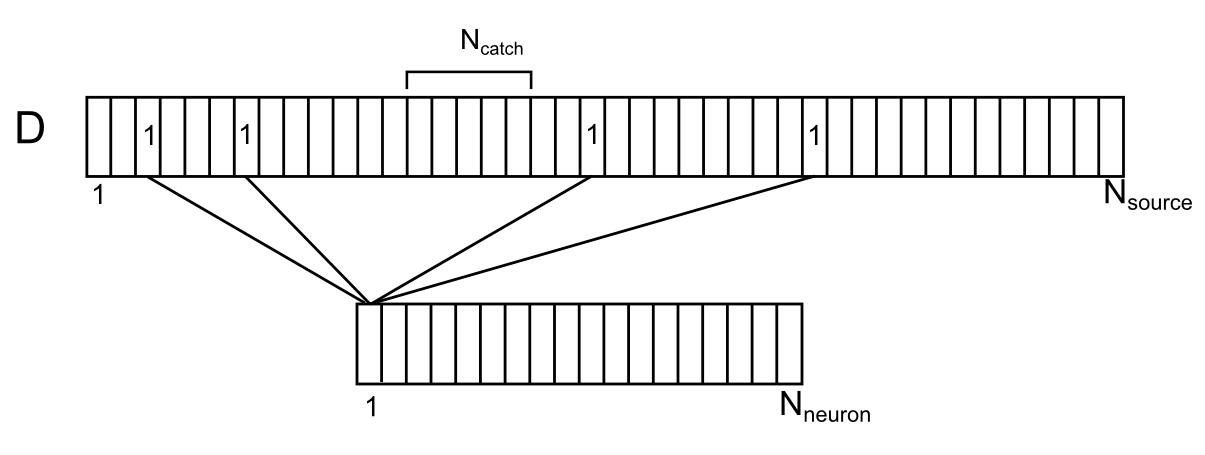
Соотнесение окружающих нейронов и их контактов на дендрите
Обозначим Nneuron – количество нейронов окружения и Nsource – количество источников для одного сегмента дендрита.
Если мы зададимся расстоянием, на которое распространяются нейромедиаторы после спиловера, то сможем определить какие синапсы способны влиять на выбранное место дендрита. Обозначим через Ncatch количество источников, способных влиять на выбранное место дендрита. Для этих источников такое место можно назвать «ловушкой».
Теперь предположим, что несколько нейронов из окружения дендрита дали спайки. Это можно трактовать как сигнал, доступный для наблюдения нашему дендритному сегменту. Обозначим Nsig – количество активных нейронов, создающих информационный сигнал. Запишем этот сигнал бинарным вектором S размерности Nsource.
Для всех позиций на дендрите, кроме самых крайних, можно определить количество активных источников (плотность сигнала), попадающих в ловушку, по формуле
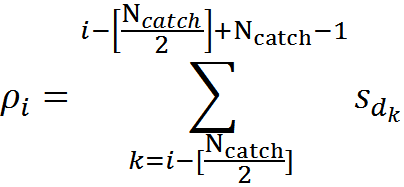
Например, для сигнала, показанного на рисунке ниже, плотность сигнала в отмеченной синаптической ловушке составит 2 (сумма сигналов от 1-го и 4-го нейронов).
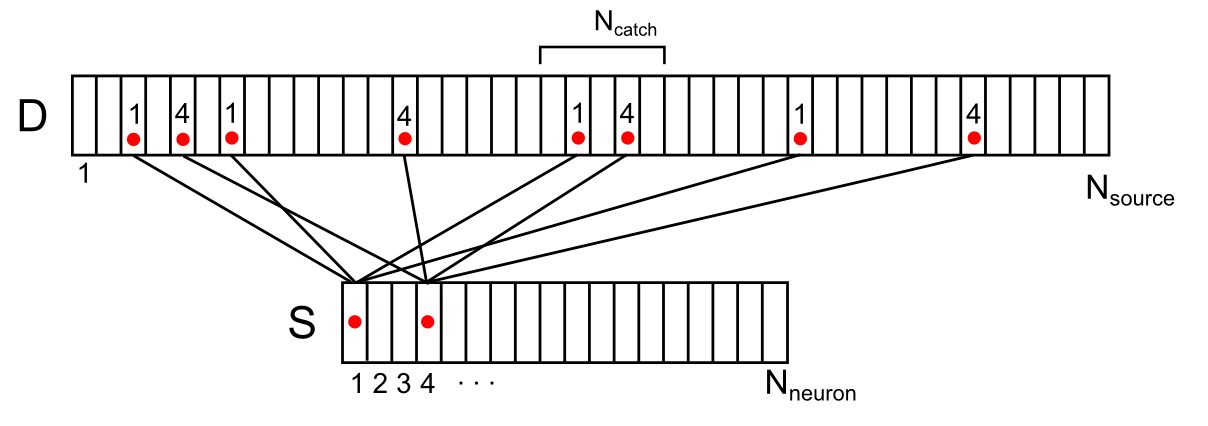
Отображение активности двух нейронов окружения на дендритный сегмент (показана только часть связей и нумерации)
Для любого произвольного сигнала можно рассчитать какое распределение плотности он создаст на дендрите. Эта плотность будет колебаться в диапазоне от 0 до Ncatch. Максимальное значение будет достигаться, когда будут активны все источники, образующие соответствующую ловушку.
Воспользуемся значениями, характерными для реальной коры крысы (Braitenberg V., Schuz A., 1998) и, исходя из них, выберем ориентировочные параметры модели:
Длина секции = 150 мкм (300 синапсов, в среднем 0.5 мкм между синапсами)
Радиус плотного ветвления дендрита = 70 мкм
Плотность нейронов в коре = 9x104/мм3
Количество нейронов в окружении дендрита (Nneuron) = 100
Количество источников для дендритного сегмента (Nsource) = 3000
Размеры ловушки (Ncatch) = 15
Будем полагать, что сигнал кодируется активностью, например, 10% нейронов, тогда
Nsig = 10
Можно рассчитать вероятность того, что для произвольного сигнала, состоящего из Nsig единиц, найдется хотя бы одно место на дендритном сегменте, где сигнальная плотность составит ровно K единиц. Для приведенных параметров вероятность принимает следующие значения:
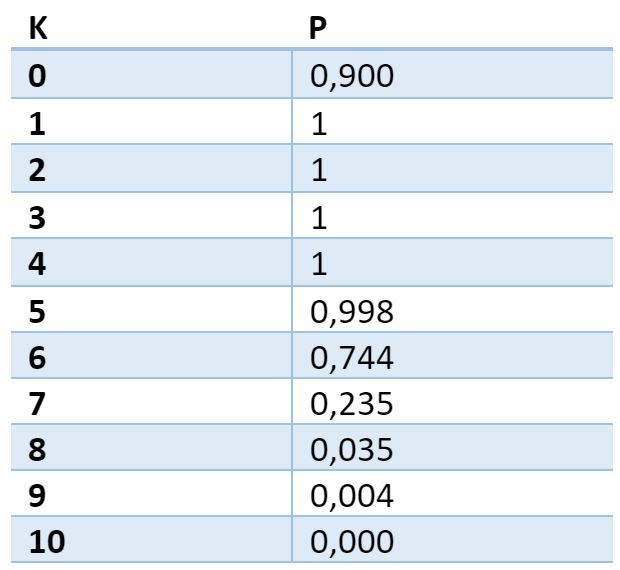
Таблица вероятности нахождения хотя бы одной ловушки с заданной плотностью. Первый столбец – требуемое количество активных источников в ловушке. Второй – вероятность нахождения на дендрите хотя бы одного места, где будет именно такое количество активных источников
Из таблицы видно, что с вероятностью близкой к 1 на любом дендритном сегменте для любого выбранного объемного сигнала найдется место, в котором сойдутся как минимум 5 аксонов активных нейронов. Это место дендрита можно считать избранным по отношению к выбранному сигналу. Если в этом месте запомнить какие именно аксоны (синапсы) были активны, то это позволит в последствии с высокой точностью детектировать повторение того-же сигнала.
Точность детектирования определяется вероятностью коллизий, то есть вероятностью того, что в каком-либо другом объемном сигнале окажутся активны те же нейроны, аксоны которых сошлись в избранном месте. То есть если, например, сигнал определяется активностью 10 нейронов из 100 и в избранном месте было зафиксировано сочетание 5 из этих 10 нейронов, то для коллизии достаточно, чтобы в каком-либо другом сигнале именно эти 5 нейронов также оказались активны.
Обозначим количество нейронов «пойманных» в ловушку, то есть активность которых совпала в избранном месте, через K. Чтобы другой сигнал, состоящий из Nsig активных нейронов, вызвал ошибочное узнавание надо, чтобы К его нейронов совпали с нейронами ловушки. Число таких совпадающих сигналов –
 . Общее количество возможных сигналов —
. Общее количество возможных сигналов — 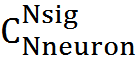 , вероятность ошибки
, вероятность ошибки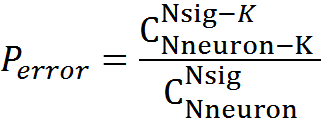
Для нашей модели при K=5 вероятность ошибки детектирования составит 3,34x10-6, при K=6, соответственно, вероятность ниже — 1,76x10-7.
Кодирование сигнала в избранном месте сочетанием нейромедиаторов
Каждое место дендрита окружено синапсами, как собственными, так и соседних дендритов. Эти синапсы являются источниками внесинаптических нейромедиаторов. Те из синапсов, что способны влиять на выбранное место дендрита образуют ловушку. Среднее количество таких синапсов для произвольного места дендрита — Ncatch. Зададимся значением K, определяющим то, сколько синапсов должно быть активно, чтобы считать место избранным по отношению к объемному сигналу. Видно, что для каждого места дендрита будет существовать достаточно много сигналов, создающих в этом месте минимум K пересечений. Чтобы мы могли с высокой точностью отследить повторение искомого сигнала необходимо не просто зафиксировать факт того, что в избранном по отношению к этому сигналу месте сработало К источников, а надо еще убедиться, что это именно те источники, что соответствуют сигналу. То есть надо по выбросу нейромедиаторов понять не просто сколько синапсов активировалось, а определить какие именно синапсы сработали на этот раз.
Как мы уже говорили, для большинства синапсов, в момент активности выделяется один «основной» нейромедиатор и в дополнение к нему один нейропептид или более ( Lundberg, J. M. 1996. Pharmacol. Rev. 48: 113-178.) (Bondy, C.A., et al. 1989. Cell. Mol. Neurobiol. 9: 427-446). Факт наличия у нейронов мозга большого количества нейромедиаторов и нейромодуляторов позволяет предположить, что основная функция такого многообразия – это создание в момент синхронной активности нейронов в каждом месте пространства уникальных сочетаний медиаторов и модуляторов. Можно предположить, что дополнительные вещества в синаптических везикулах распределены по синапсам так, чтобы обеспечивать в каждом месте пространства максимально разнообразие распределение. Если это так, то детектирование определенного сочетания активности синапсов сводится к определению соответствующего этим синапсам уникального набора испускаемых веществ.
Таким образом, если в избранном по отношению к определенному сигналу месте дендрита поместить детектор, чувствительный к сочетанию веществ, характерному для этого сигнала, то срабатывание этого детектора будет с очень высокой вероятностью говорить о повторении исходного сигнала.
Теперь мы знаем, что на каждой дендритной веточке всегда найдется место, которое окажется избранным по отношению к любому сигналу окружающих нейронов. Нам осталось понять, как веточка дендрита может запомнить, что это ее избранное место должно реагировать на определенное сочетание нейромедиаторов.
Рецепторы нейронов, как элементы памяти
Описывая работу нейрона, мы говорили, что окружающие вещества влияют на работу нейрона через его рецепторы. Рецепторы бывают ионотропные и метаботропные. Ионотропные рецепторы связываются с нейромедиаторами, высвобождающимися в синапсе, что вызывает изменение их конформации. Конформация молекулы – это изменение ее пространственной структуры без изменения самого состава молекулы.
Ионотропные рецепторы одновременно являются ионными каналами. Изменение конформации открывает ионный канал рецептора, что приводит к перемещению ионов и, соответственно, изменению мембранного потенциала.
Метаботропные рецепторы не имеют ионных каналов и действуют иначе. Той частью, что находится внутри нейрона они связаны с, так называемыми, G-белками. Когда эти рецепторы взаимодействуют со своими сигнальными веществами их конформация меняется и G-белок отпускается. Это ведет к разным возможным последствиям. Одно из возможных последствий – это открытие G-белками соседних ионных каналов, которые достаточно быстро меняют локальный мембранный потенциал нейрона. Что, в свою очередь, вызывает токи в дендрите и может стать причиной дендритного спайка.
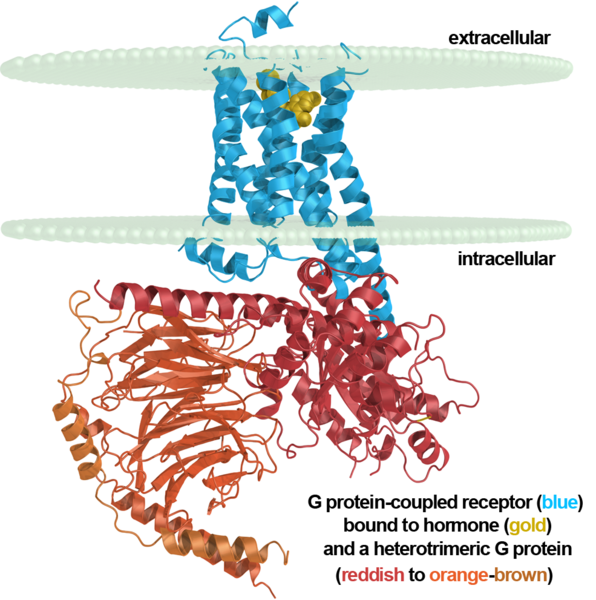
Метаботропный рецептор, мембрана нейрона и G-белок
Метаботропные рецепторы, в основном, располагаются вне синапсов и являются мишенями именно для внесинаптических нейромедиаторов. Рецепторы, в основном, объединяются в кластеры и действуют совместно. Кластеры метаботропных рецепторов, по сути, являются разнообразными детекторами, настроенными на определенные сочетания нейромедиаторов.
Кластеры метаботропных рецепторов очень подходят на роль элементов энграммы для нашей модели. Рядом с каждым синапсом могут находиться сотни таких кластеров. В них заранее случайным образом составлены сочетания рецепторов, потенциально чувствительные ко многим возможным в этом месте комбинациям внесинаптических нейромедиаторов, то есть таких нейромедиаторов, которые могут выделиться во внешнюю среду из соседних синапсов.
То есть расположенные в большом количестве в каждом месте дендрита метаботропные рецепторы могут являться «заготовками» для будущих энграмм. Переход «заготовок» в энграммы можно описать так. Предположим, что кластеры рецепторов, относящиеся к какой-либо дендритной ветке, изначально неактивны и никак не влияют на ее работу. Когда этой ветке требует произвести запоминание он дает знать об этом всем метаботропным рецепторам. Таким сигналом, например, может быть небольшая общая деполяризация мембраны этой ветки. Как мы говорили выше, где-то на этой ветке обязательно найдется избранное место. То есть место, где рядом с этой веточкой окажется сразу несколько активных синапсов. Если в этом месте веточки найдется кластер с рецепторами, чувствительность которого совпадет с коктейлем нейромедиаторов, то этот кластер должен будет перейти в активное состояние и впоследствии всегда реагировать на появление своего коктейля. Если до этого кластер не влиял на работу веточки дендрита, то теперь он должен будет при появлении своего коктейля создавать возбуждающий постсинаптический потенциал.
Оказывается, что метаботропные рецепторы, действительно умеют так себя вести. Определенные обстоятельства могут погружать наружные и внутренние части рецептора встречно в мембрану, лишая рецептор чувствительности. Или, наоборот, чувствительные части рецептора могут быть вытолкнуты из мембраны. В таком чувствительном состоянии рецептор может пребывать какое-то время, затем он может вернуться в исходное состояние – это соответствует краткосрочной памяти. Рецептор может и надолго зафиксировать такое чувствительное состояние. Если складываются подходящие условия, то начинаются процессы адгезии и полимеризации, которые могут оставить рецептор в чувствительном состоянии на дни и недели. Если процесс фиксации, который длится, предположительно, около месяца не прервется, то состояние рецептора зафиксируется навечно или правильнее сказать пожизненно. Все это соответствует различным стадиям консолидации долгосрочной памяти.
Механизмы, которые управляют таким поведением метаботропных рецепторов подробно изучил и описал А. Н. Радченко (Информационные механизмы мозга, 2007). Кстати, Радченко первый высказал предположение, что именно кластеры метаботропных рецепторов с их конформационными переходами и есть элементы энграмм.
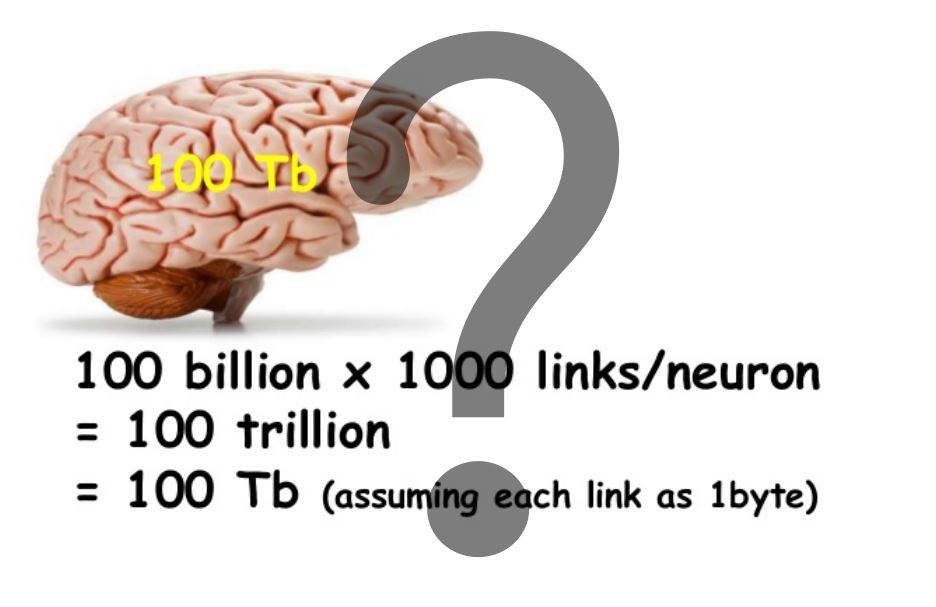
Для памяти, основанной на пластичности синапсов, емкость считается достаточно просто. Пример такого расчета приведен на заглавной картинке. Заметим, что в нашей модели емкость памяти на дендритах, где-то в 1000 раз больше. И это еще не все.
В той пространственной структуре, что создается переплетением аксонов и дендритов работает идеология «избранных мест». То есть, чтобы рецепторы “были в деле” им необязательно относиться к тому дендриту, которому принадлежит синапс, через который передается сигнал. За счет того, что нейромедиаторы распространяются в межклеточном пространстве, “в деле” могут быть любые рецепторы, просто, геометрически находящиеся поблизости. Причем это, вообще, не обязательно должны быть рецепторы, принадлежащие именно нейронам.
Так, глиальные клетки коры, плазматические астроциты (рисунок ниже), имеют наборы тех же рецепторов, что и нейроны, а значит могут быть участниками механизмов памяти.
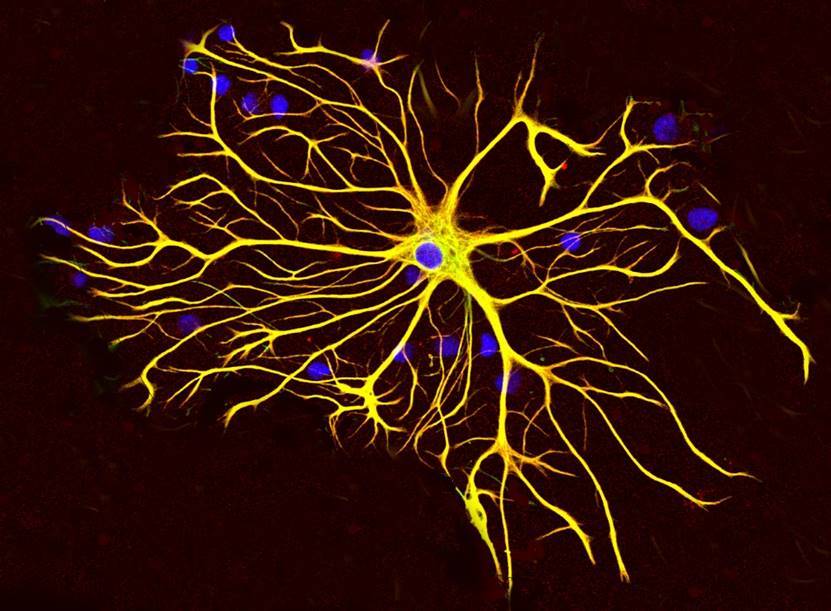
Плазматический астроцит
В коре количество астроцитов превышает количество нейронов. Астроциты коры имеют короткие ветвящиеся отростки. Этими отростками они охватывают находящиеся поблизости синапсы (рисунок ниже).
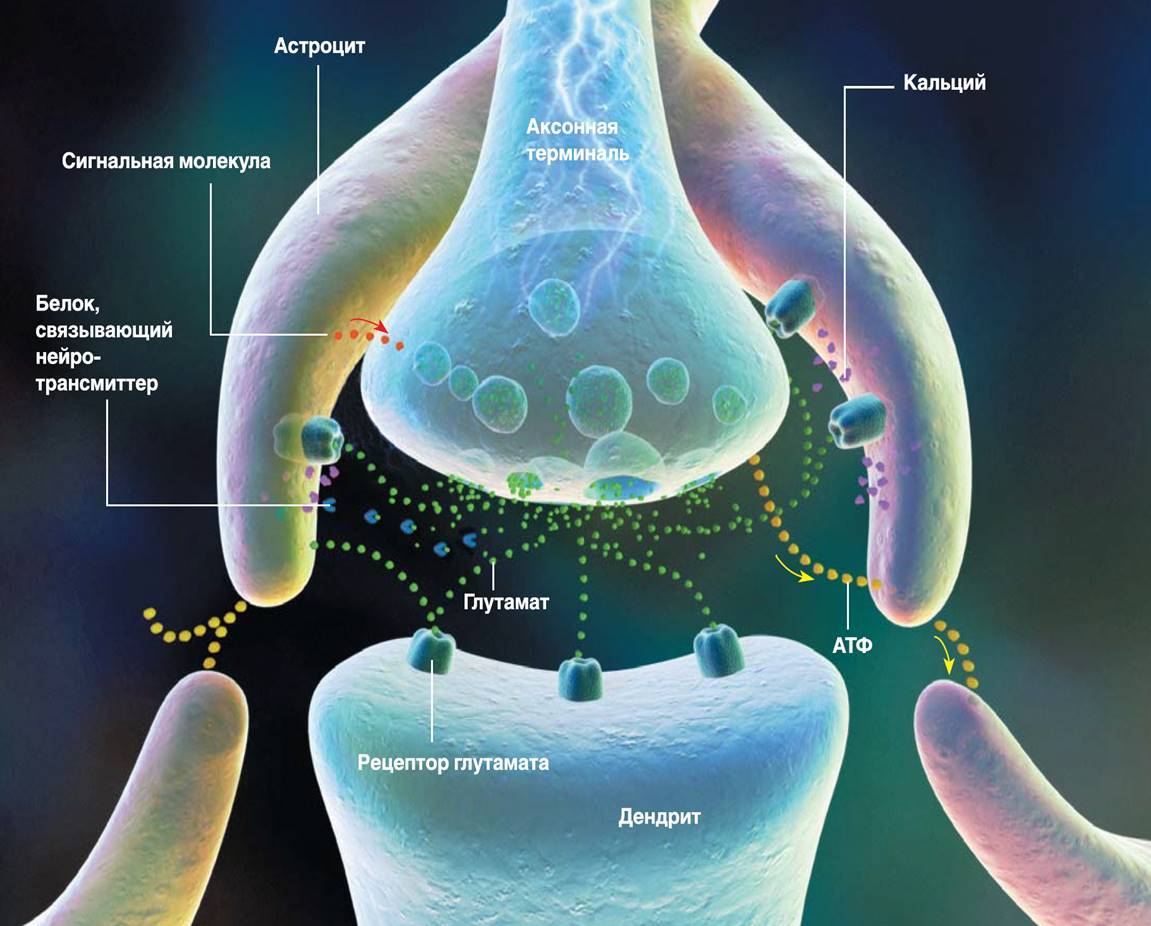
Трехсторонний синапс (R. D. Fields, B. Stevens-Graham, 2002)
Астроциты способны как усиливать реакцию синапса за счет выброса соответствующего медиатора, так и ослаблять ее за счет его поглощения или выброса связывающих нейромедиатор белков. Кроме того, астроциты способны выделять сигнальные молекулы, регулирующие выброс нейромедиатора аксоном. Концепция передачи сигналов между нейронами, учитывающая влияние астроцитов, называется трехсторонним синапсом (R. D. Fields, B. Stevens-Graham, 2002). Возможно, что трехсторонний синапс – это основной элемент, реализующий механизмы взаимной работы различных систем памяти.
Роль гиппокампа. Информация в идентификаторах. Кольцевые идентификаторы
В описываемой модели памяти чтобы на какой-либо зоне коры могли формироваться воспоминания кроме самой информационной картины на нее должны подаваться сигналы идентификаторов воспоминаний. Так как зоны коры выполняют различные функции, то уместно предположить, что и идентификаторы воспоминаний различаются для разных зон или групп зон.
Некоторые известные участки мозга в силу своей специфики хорошо подходят на роль источников идентификаторов. Так, верхние бугорки четверохолмия могут быть источником идентификаторов для зрительных зон. Нижние бугорки четверохолмия подходят на роль генераторов идентификаторов для слуховых зон коры. Наиболее показательный орган, связанный с памятью, – это гиппокамп, который хорошо подходит на роль генератора ключей воспоминаний для зон префронтальной коры.
В 1953 году пациенту, которого принято называть H.M. (Henry Molaison), при попытке вылечить эпилепсию было произведено двустороннее удаление гиппокампа (W. Scoviille, B. Milner, 1957). В результате у H.M. полностью исчезла способность что-либо запоминать. Он помнил все, что было с ним до операции, но что-либо новое вылетало у него из головы, как только переключалось его внимание. Кто смотрел фильм «Помни» («Memento») Кристофера Нолана, тот хорошо поймет, о чем разговор.

Henry Molaison
Случай H.M. достаточно уникален. В других случаях, связанных с удалением гиппокампа, где не было такого полного двустороннего разрушения, как у H.M., нарушения памяти либо были не столь ярко выражены, либо отсутствовали вовсе (W. Scoviille, B. Milner, 1957).
Полное удаление гиппокампа делает невозможным формирование новых воспоминаний. Нарушения в работе гиппокампа могут привести к синдрому Корсакова, который так же сводится к невозможности фиксировать текущие события, при сохранении старой памяти.
Достаточно распространенное представление о роли гиппокампа сводится к тому, что гиппокамп – это место хранение текущих воспоминаний, которые впоследствии перераспределяются по пространству коры. В описываемой модели роль гиппокампа иная, он создает уникальные ключи воспоминаний.
Созданные гиппокампом ключи через систему проекций распространяются на соответствующие зоны коры. Интерференция идентификаторов гиппокампа и информационных описаний создает память. При этом память сразу образуется «на своем месте» и не перемещается между гиппокампом и корой. Такое представление хорошо согласуется с экспериментальными данными. Действительно, удаление гиппокампа делает невозможным новое запоминание, так как исчезает генератор ключей воспоминаний. Старые воспоминания остаются нетронутыми, так как уже не зависят от гиппокампа. Их идентификаторы могут быть извлечены и использованы без задействования гиппокампа.
Но главные доводы в пользу предлагаемой роли гиппокампа связаны с функциями, которые обнаружены в гиппокампе и на первый взгляд не имеют прямого отношения к механизму памяти.
В 1971 году Джон О‘Киф открыл в гиппокампе клетки места (O’Keefe J., Dostrovsky J., 1971). Эти клетки реагируют подобно внутреннему навигатору. Если крысу поместить в длинный коридор, то по активности определенных клеток можно будет точно сказать, в каком его месте она находится. Причем реакция этих клеток не будет зависеть от того, как она попала в это место.
В 2005 году в гиппокампе были обнаружены нейроны, кодирующие положение в пространстве, образующие нечто наподобие координатной сетки (Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I., 2005).
В 2011 году оказалось, что в гиппокампе есть клетки, которые определенным образом кодируют временные интервалы. Их активность образует ритмические узоры, даже если ничего другого вокруг не происходит (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011).
Хранение данных в виде пар «ключ-значение» создает ассоциативный массив. В ассоциативном массиве ключ имеет двойную функцию. С одной стороны, он является уникальным идентификатором, который позволяет отличить одну пару от другой, с другой стороны, сам ключ может нести информацию сильно облегчающую поиск. Например, файловая система компьютера может рассматриваться как ассоциативный массив. Значение – это информация, хранящаяся в файле, ключ – информация о файле. Информация о файле – это путь, указывающий на место хранения, имя файла, дата создания. Для фотографий дополнительна информация – геотеги, координаты места, где сделан снимок. Для музыкальных файлов – название альбома и имя исполнителя. Все эти данные о файлах образуют составные сложные ключи, которые не только однозначно идентифицируют файлы, но и позволяют вести поиск по любому из полей ключа или любому их сочетанию. Чем подробнее составлен ключ, тем гибче оказываются возможности поиска.
Так как мозг реализует те же информационные задачи, что и компьютерные системы, то логично предположить, что хранение мозгом данных в виде пар «ключ-значение» будет сопровождаться создание ключей наиболее удобных для поиска. Для воспоминаний, с которыми имеет дело человек разумный набор дескрипторов ключа должен включать в себя:
- Указание места действия;
- Указание положения в пространстве;
- Указание времени события;
- Набор понятий, советующих основному смыслу происходящего. Некий аналог ключевых слов, описывающих содержание статьи.
Очень похоже, что гиппокамп не просто работает с местом, положением в пространстве и временем, а использует эти данные именно для составления сложных информационных ключей воспоминаний. По крайней мере, это очень хорошо объясняет, почему столь разнообразные функции сошлись в одном месте. Причем месте, непосредственно отвечающем, за формирование памяти.
Особый интерес представляет временное кодирование. Человеческая память не просто позволяет вспоминать статические картины, она способна воспроизводить последовательность сцен с сохранением их хронологии. Соответственно, в систему кодирования памяти должна быть заложена такая возможность. Было показано, что в гиппокампе есть клетки времени, создающие ритмические узоры (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011). Цикличность узоров наводит на мысль, что гиппокамп может использовать для создания временных полей идентификаторов событий те же приемы, что использует и человек для измерения времени.
Чтобы описать ход времени мы используем часы и календарь. В основе и того и другого лежат кольцевые идентификаторы. Минута состоит из 60 секунд. Это значит, что 60 идентификаторов последовательно сменяют друг друга, причем после 60 секунды опять следует первая. Аналогично с минутами в часе, часами в сутках, днями в месяцах, днями в неделях, месяцами в годах, годах в столетиях. То есть несколько кольцевых идентификаторов с разной периодичностью позволяют идентифицировать любой момент времени.

Похоже, что гиппокамп использует схожую систему временных кольцевых идентификаторов, что, собственно, и наблюдалось в опытах. Но, что особенно интересно, что такая система позволяет не только идентифицировать воспоминания, но и воспроизводить их последовательность. Если мы знаем правила следования идентификаторов друг за другом, то имея идентификатор одного события мы легко можем получить идентификатор события, следующего хронологически за ним и так далее.
Надо отметить, что кольцевые идентификаторы удобны не только для описания времени, но и для многих других ситуаций.
То, что мы описали в этой части, очень важно для понимания работы мозга, но не содержит, пока, никакой «магии». Да, это показывает где может находиться память и в каком виде она может храниться, но это не говорит ничего о том, как же мозг с легкостью решает задачи, непосильные компьютеру. Компьютер, конечно, может считать быстрее человека, но зато человек способен понимать смысл происходящего. В следующей части я расскажу, что вообще есть «смысл» и постепенно, станет понятно зачем нам так долго пришлось описывать работу нейронов и механизмы голографической памяти.
Алексей Редозубов
Логика сознания. Вступление
Логика сознания. Часть 1. Волны в клеточном автомате
Логика сознания. Часть 2. Дендритные волны
Логика сознания. Часть 3. Голографическая память в клеточном автомате
Логика сознания. Часть 4. Секрет памяти мозга
Логика сознания. Часть 5. Смысловой подход к анализу информации
Комментарии (39)

AlekseiMorozov19730316Ru
07.09.2016 00:37>… оказывается очень трудной задачей увязать конкретный нейрон,
>если приписывать ему функции детектора какого-либо свойства,
>и память, которая в силу многих причин,
>не должна быть жестко привязана к определенному нейрону…
«Увязка» нюансов распознавания и ассоциирования является, безусловно, «трудной», но всё-таки разрешимой «задачей».

AlexeyR
07.09.2016 00:41Тут несколько о другом. Если детектор некого свойства привязан к нейрону, то тогда и память, содержащая это свойство, должна быть где-то поблизости от него. И как-то на него опираться. Это создает техническую сложность, которую практически не преодолеть.
AlekseiMorozov19730316Ru
07.09.2016 09:09Отнюдь. Что память «должна быть где-то поблизости от него» [нейрона] — заблуждение. Широковещательное распространение паттернов нейронов мозга преодолевает эту сложность посредством гибкого ассоциирования выходных паттернов одних нейронов с входными паттернами других нейронов. По сути, входной и выходной паттерны нейрона — это и есть его память. Она «не поблизости», она — он и есть. Память — это сам нейрон.
AlexeyR
07.09.2016 14:43Если памть сам нейрон, то количество воспоминаний равно количеству нейронов? Если удалить нейрон исчезнет воспоминание? Память о каждом событии есть везде или в конкретных местах?

napa3um
07.09.2016 14:56Можно допустить формулировку, что совокупность всех состояний нейронов представляет собой суперпозицию всех «воспоминаний» человека. Удаление нейрона увеличивает _дискретность_ этих воспоминаний (незначительно уменьшает вероятность отличить одно воспоминание от другого). Вполне удобная интерпретация, на мой взгляд (и не противоречит вашей модели, между прочим).
AlexeyR
07.09.2016 15:07Сложности возникают в момент реализации. Мы ограничены биологией нейронов. Далеко не все идеи удается правдоподобно сопоставить с реальными клетками.
napa3um
07.09.2016 15:10Такая формулировка не противоречит вашей модели (формулировка вообще очень абстрактна и является, по-сути, неявным определением термина «воспоминание» как состояния мозга, различимо влияющего на поведение человека — например, его признания о том, что он что-то вспомнил). А ваша модель, если верить вам, как раз хорошо сопоставляется с биологией.
napa3um
07.09.2016 15:23(Грубо говоря, мы просто отказываемся от формулирования того, как нейроны хранят свои знания, мы заталкиваем их в чёрный ящик и даём оценку «сверху» о том, что они не смогут запомнить больше, чем сумма количества всех возможных состояний всех нейронов. Важно понимать, что определение «количества состояний» уже требует привлечения формализации среды, в которой существует такой чёрный ящик и для которой состояния этого чёрного ящика вообще могут быть различимы — это и непосредственно прилагаемое к мозгу тело, и состояние пространства вокруг тела, и даже социальные контрагенты — т.е., различимость состояний мозга всегда относительна задачи исследователя и выбранного «масштаба» измерений. Наверное, весь этот косноязычный текст можно свести к ссылке на описание агентного моделирования. Кстати, если выбрать в качестве агента не мозг, а нейрон, то можно увидеть и некоторые переоткрытые вами принципы распространения автоволн в многоагентной среде.)
napa3um
07.09.2016 15:44(«Чем сумма количества всех возможных состояний всех нейронов» — не сумма, а произведение, конечно.)
AlexeyR
07.09.2016 16:39Автоволны в многоагентной среде — это, все-таки, больше волны. То есть некие периодические процессы. В моей модели мы имеем дело с распространение уникальных узоров, что совсем другое.
Если не усложнять, то я пытаюсь показать, что каждое место мозга может хранить пару ключ-значение без привязки к конкретным нейронам. Функции нейронов получаются не более чем совместное воспроизведение кода, непривязанного к конкретному месту.
А так же, я показываю, что каждый небольшой участок коры может хранить достаточно много таких пар «ключ — значение».napa3um
07.09.2016 17:01Мне кажется, что я понимаю ваши интерпретации, и мне кажется, что они эквивалентны сказанному мной (в моих формулировках тоже нейроны — это субстрат, проявляющий свои _важные_ свойства как целое; отдельный нейрон не хранит осмысленные «воспоминания», но «вкладывается» «количественно» в свойства субстрата, хранящего такие воспоминания; а автоволны в широком смысле — это распространение межэлементных взаимодействий по субстрату из любых активных элементов, в том числе и нейронов, вне зависимости от их регулярности и ритмичности). Но не буду мутить воду и навязывать свои термины и интерпретации, дождусь следующих частей.
AlekseiMorozov19730316Ru
07.09.2016 15:14>Если память сам нейрон,
>то количество воспоминаний равно количеству нейронов?
Нет, разумеется. Слишком абсурдное упрощение. Любое воспоминание «крошится» в «пыль» паттернов. И уже эти паттерны мало по малу у нейронов изменяются. Память — вообще сродни иллюзии.

Halt
07.09.2016 07:18+2Спасибо за статью! Модель действительно выглядит крайне любопытно.
Описанные вами механизмы формирования воспоминаний вполне согласуются с жизненным опытом и прочими «техниками запоминания». Считается, что запомнить что-то тем проще, чем больше разных органов чувств и эмоций будет вовлечено в процесс. Насколько я понимаю, это хорошо согласуется с вашей моделью: чем больше базовых идентификаторов окажется в составном ключе (время, место, положение и т.д.), тем проще будет потом выудить записанную информацию, что на бытовом уровне мы воспринимаем, как «хорошо/плохо запомнил». То есть, проблема не в том, чтобы записать информацию, а в том, чтобы подобрать такие ключи, по которым будет легко вспомнить.
Вопрос: кто в итоге решает, что? надо запомнить, а что нет? Вы пишете, что кластер будет запоминать активность, если он уже? находится в деполяризованном состоянии. Кто его деполяризует? Предполагается, что продолжительная/повторяющаяся активность нейронов повышает шансы того, что кластеры «сработают» и тем самым отразят факт того, что в данной точке пространства такие волны уже были?
Второй момент из личного опыта: когда я стараюсь вспомнить некоторую хорошо известную мне локацию X, то практически непроизвольно в памяти начинают всплывать картины из ближайшего окружения X. Заметил, что помимо окружающих картин, опять же непроизвольно возникают образы локаций, находящихся на пути из моей текущей точки Y в точку X.
Если я правильно понимаю, это вполне можно описать последовательностью воспоминаний, где каждое следующее вылавливается по ключу места текущего. В сумме получается что-то вроде механизм поиска маршрута из одного известного места в другое. Действительно, механизм такого поиска очень напоминает алгоритмы прокладки маршрутов: Дейкстры или A* в зависимости от того, в каком пространстве лежат воспоминания.
Поиск пути по воспоминаниям «первого уровня», привязанным к геометрии местности больше напоминает А*, тогда как поиск пути в абстрактном графе (выбор станций в метро) больше похож на Дейкстру. Разумеется, если точки находятся в разных анклавах и субъекту неизвестен путь из одной точки в другую, то никаких промежуточных ассоциаций не возникает в принципе.
Вопрос: известны ли вам исследования на эту тему? Да, я читал про нейроны положения у крыс в лабиринте. Меня интересует именно активность при поиске пути. На базе вашей теории можно сделать вполне конкретные прогнозы относительно того, какого рода активность должна возникать.
Volutar
07.09.2016 14:46+3Стоит упомянуть один нюанс связанный с пациентом H.M. (у Алексея в статье про это не упомянуто). После удаления гиппокампа старые воспоминания у него не остались в том виде, котором были до этого. Все старые воспоминания (насколько я помню) «рассыпались, словно фрагменты паззла по комнате». Т.е. помнятся отдельные факты, но не помнятся с чем и как он связаны, ни контекст, ни обстоятельства их получения, ни время. Для этих былых фактов остаётся только ассоциативная связность через другие области мозга.
Есть все признаки того, что гиппокамп генерирует хронологически связанные идентификаторы (в терминах AlexeyR), которые привязываются к энграммам «внешних ощущений», идущих с первичных зон через инсулярную кору (в которой происходит интеграция всех сенсорных видов-модальностей в общую картину самоощущения). Связанность ощущений себя в том или ином месте сама по себе создаёт топологическую структуру-карту в гиппокампе — процесс вспоминания по сути активирует зоны «аналогичных ощущений», которые, способом, действительно похожим на Дейкстру (волновая реактивация), имеют доступ до соседних позиций. При этом пока положение до конца не распознано — в гиппокампе продолжают «отвечать» несколько позиций, имеющих сходства, пока в определённый момент «неоправданные гипотезы» не будут подавлены при противоречивости наблюдаемого с воспоминаниями.
При этом нет никакой общей глобальной карты. Карты все достаточно локальны, т.е. ограничиваются областью одновременно обозреваемого. Свежая информация на сей счёт. Глобализация ориентации происходит уже при достаточно глубоком анализе и связывании областей через лобные доли.
Т.е. всё достаточно красиво вписывается в эту концепцию. Стоит упомянуть любопытный момент в гиппокампе — это так называемые «клетки скорости». Точнее не столько клетки, сколько определённая активность в определённом месте, не зависящая от положения в пространстве, а зависящая только от скорости. Ну, и, логично, что никакого спидометра нету, и скорость эта тоже берётся от собственных ощущений.AlexeyR
07.09.2016 15:03+2Volutar, спасибо за важны дополнение. Я боюсь, что статьи и так сильно перегружены информацией и стараюсь писать только самое главное. Относительно HM. Чтобы вспомнить что-то фрагментарно достаточно по ассоциациям найти идентификатор воспоминания и восстановить связанную с ним картину. Но что-бы перейти к последовательному хронологическому воспоминанию надо создать следующий по порядку хронологический идентификатор. В стать описаны кольцевые идентификаторы. Для них это означает, что надо надо сначала выставить их в соответсвии с воспоминанием, а затем в зависимости от того какой мы выбрали временной шаг воспоминания перевести один из них на шаг вперед или назад. Это выглядит, как перевод одной из стрелок часов. Знания о том, как упорядочены кольцевые идентификаторы времени содержится в гиппокампе. Он и генерирует эти последовательности. Удаление гиппокампа делает невозможным последовательный «перевод стрелок».
Halt
07.09.2016 15:49При этом нет никакой общей глобальной карты. Карты все достаточно локальны, т.е. ограничиваются областью одновременно обозреваемого. Свежая информация на сей счёт. Глобализация ориентации происходит уже при достаточно глубоком анализе и связывании областей через лобные доли.
Тем не менее, это хорошо заметно, когда бродишь в незнакомом городе. Сначала ориентируешься только в крайне ограниченных областях пространства: аэропорт/метро/гостиница/квартира друзей/достопримечательность X. По мере того, как начинаешь активно ходить пешком и внезапно выходишь в знакомое место, анклавы начинают объединяться в сознании в регионы большего размера.
До сих помню свое удивление, когда гуляя по Питеру, вышел в знакомое место, в которое раньше добирался только через метро, а на самом деле оно оказалось совсем рядом. Прямо физически ощутил, как в голове «шестеренки крутятся» и карта достраивается. Что характерно, после этого начинаешь адекватно оценивать расстояния до точек внутри известной области.
AlexeyR
07.09.2016 14:51+1Halt, вы очень здраво рассуждаете. Но проще будет, когда картина увидится более полно. Сейчас я описал сам механизм, что позволяет создавать и извлекать воспоминания. Что управляет извлечением воспоминаний и каковы законы этого будет описано позже. Там все очень интересно и не сводится к жестким закономерностям типа: после этого всега следует то.
Halt
08.09.2016 10:44Понятно. Ну и еще момент вдогонку, ответ на который хотелось бы услышать в будущем.
Как определяются границы распространения волн (в пространстве и времени)? Интуитивно ясно, что если каждая волна от каждого ключа будет развиваться неподконтрольно, то ничего хорошего не выйдет. Доводя до абсурда: каждый раз, когда мы заходим к себе домой или на работу, нас бы заваливало ворохом воспоминаний, связанных с этим местом.napa3um
08.09.2016 11:38Распространение волн происходит в активной среде из нейронов, которая как бы оказывает им сопротивление тем, что ресурс нейрона на формирование спайка ограничен и нейрону нужно время на восполнение этих ресурсов. И все взаимопересекающиеся волны, получается, конкурируют за эти ресурсы для спайков. Это примерно как волны прибоя на песчаном пляже — вроде и проходят друг через друга, не задевая, но и не складываются бесконечно «стопкой», вода постоянно утекает в море так, что на песке можно увидеть фронт одновременно всего двух-трёх волн.
Halt
08.09.2016 11:42На интуитивно-бытовом уровне я это понимаю. Интересует более формальный подход к проблеме, поскольку скорее всего, все гораздо сложнее и интереснее, чем аттенюация сигнала.

Heath
07.09.2016 08:42Алексей, спасибо! Очень круто и ждём про смыслы! Особенно круто, если получится увязать с тем, как смыслы передаются (формируются) от мозга к мозгу. И насколько вообще правильно говорить о том, что смыслы могут совпадать у разных людей. А также о том, что такое рефлексия и внутренний наблюдатель, перед которым эти смыслы разворачиваются.
AlexeyR
07.09.2016 15:10+1Постараюсь не разочаровать. Про передачу смысла и его совпадение или несовпадение ответ будет полный. Про внутреннего наблюдателя будут гипотезы, которые, возможно, заставят посмотреть на это с неожиданной стороны.

BalinTomsk
07.09.2016 10:34--но что-либо новое вылетало у него из головы, как только переключалось его внимание
Мне кажется это у всех приходит с возрастом. Что сейчас предлагает современная наука для решения этой проблемы?
Учу японский -совершенно не ложится в голову.
Была новость пару лет назад что при повторении слова 124 раза образуется устойчивое (на всю жизнь) соединение между нейронами. Мне кажется врут.
Istizar
07.09.2016 15:11На Coursera есть курс Learning How to Learn. Там много чего интересного о процессе обучения. Например, повторение 124 раза одного слова ничего устойчивого создать не может, если повторять подряд. Повторения должны быть разбросаны во времени, например одни раз в день, несколько дней в неделю. Если это будет происходить в разных местах, то эффект будет лучше (не будет привязки к определенной локации). В процессе обучения вообще плохо биться головой о стену, важно периодически переключаться из сфокусированного режима (focused) в рассеянный (diffused) и делать паузы для отдыха. Да много чего еще.
Там же речь шла об исследования, подтверждающих, что в любом возрасте в определенной области мозга, отвечающей за обучение, продолжает генерация новых клеток. Это область — гиппокамп, генерирующий ключи, которые образуют память в других областях мозга в предлагаемой выше модели.
AlekseiMorozov19730316Ru
07.09.2016 12:05-1>… до сих пор, как ни удивительно это звучит,
>нет ни одной теории памяти.
>То есть, есть разные предположения о том,
>что может быть связано с памятью…
Обособлять «теорию памяти» нет смысла. Мозг — это не компьютер с процессором и памятью. Знания о нейроне — и есть теория памяти. Ассоциирование нейронов посредством широковещательного распространения паттернов нейронов мозга устраняет сложность понимания в вопросах работы памяти.
Introducer
07.09.2016 15:13+1>>Для воспоминаний, с которыми имеет дело человек разумный набор дескрипторов ключа должен включать в себя:
Должен включать в себя контекст :)
Про кольцевые идентификаторы времени… Думаю что у меня они такие же как и кольцевые идентификаторы для Byte (0..255). А ещё точней, для позиционной системы исчисления с разными размерами разрядов. Есть порядок, есть больше, есть меньше. Подозреваю что у других людей как-то иначе. Т.е. у меня это не столько про время, сколько про систему кодирования для чисел. Время потому что в числах :)
Ещё кстати есть такая интересная тема, как память о воспоминаниях. Я могу наблюдать за тем как я думаю, или наблюдать за своим внутренним диалогом, анализировать его, потом анализировать свой анализ :) Т.е. есть память не только о внешних событиях, но и о внутренних, и память об анализе памяти (а анализ есть целеноправленное направление внимания на определённое место памяти (что есть внимание и как оно навигируется в место, на каком языке оно получает команды, и почему ему определённо нужно именно туда и именно сейчас — нужно думать)). Причём память не точная, и автоматически возникающие интерпретации зависят от того в каком состоянии вспоминаешь и в каком контексте, для каких целей (важны ли они, интересны ли). Причем часто интерпретации прицепляются к тому месту памяти и доступны при последующих просмотрах, а некоторые можно отбросить как неверные (противоречивые интерпретации обычно не уживаются, но старые помечаются как неверные или забываются (но похоже они всё-равно доступны при потребности)).
Ну и ещё эмоции могут прицепиться, или ощущения (вспоминание об апельсине в состоянии голода может вызвать слюновыделение и вкус апельсина во рту).
И ещё бывают сновидения с прекрасными и не очень мирами, и бывает даже память о них. В некоторых работает критическое мышление. В некоторых избирательно.
Я бы сказал что можно не привязываться к внешнему миру для понимания работы памяти. Всё то же есть и в мирах сновидений. И гравитация, и свет, и звуки, и эмоции, и боль, и память, и забывание, и время, и причины со следствиями. Правда осознанность там есть не всегда. Но она и тут у многих не всегда есть :)Volutar
07.09.2016 15:50Память о воспоминаниях — это уже часть концепции «теории разума», когда в том числе и собственная личность воспринимается как «внешняя». В рамках каждой модели личности есть некий архив сведений, в том числе и хронология получения сведений. Правда, там всё достаточно грубо — мало кто из людей выстраивает подробные модели других людей. Собственная модель — само собой очень подробная (т.к. имеется полный доступ до _всего_). Различные виды психических отклонений, включающих в себя диссоциацию личности (диссоциативная фуга, диссоциативное расстройство идентичности) — очень хорошие иллюстрации «глюков» этого механизма.

Shamov
07.09.2016 17:33Мне особенно нравится идея с хэш-функциями. Она полностью объясняет феномен дежавю, от которого у меня всё время мурашки. Прям ужас охватывает. Умом понимаешь, что эту конкретную сцену, которая прямо сейчас перед глазами, ты раньше видеть никак не мог. Это абсолютно исключено. Но, тем не менее, она почему-то выглядит очень знакомо. Вплоть до мельчайших деталей. Коллизия при расчёте хэша замечательно объясняет эту, как бы, бредовую ситуацию.
vagran
08.09.2016 21:59Отсюда интересное следствие — чем чаще возникает ощущение дежавю, тем больше вероятность, что место в «хеш-таблице» начинает заканчиваться, capacity мозга исчерпан.
shabanovd
08.09.2016 22:39Возможно, что это компенсируется ростом, т.е. увеличением объема, что уменьшает коллизии.
AlekseiMorozov19730316Ru
07.09.2016 18:37>… Для памяти, основанной на пластичности синапсов,
> емкость считается достаточно просто.
> Пример такого расчета приведен на заглавной картинке.
> Заметим, что в нашей модели емкость памяти на дендритах,
> где-то в 1000 раз больше…
Если это разные типы моделей, то сравнение «емкостей памяти» некорректно.
napa3um
А вы пытались реконструировать эволюционную историю, которая бы привела к такому механизму сознания (имеются ввиду описанные во всех статьях цикла, не только в данной)? Можно ли «отмотать» историю от мозга человека с описываемыми вами механизмами до, скажем, нескольких сотен тысяч нейронов без особых дифференциаций (до так называемой диффузной нервной системы) так, чтобы приспособленность организма (связанная с функционированием мозга) получалась бы монотонно убывающей «в прошлое»? Есть хотя бы наброски подобных рассуждений, или эволюционная обоснованность механизмов мозга как критерий достоверности модели вами пока не рассматривается?
AlexeyR
Очень хороший вопрос. У меня есть очень сильные доводы в пользу этой модели, основанные именно на эволюционном подходе. Но эти доводы настолько интересны, что позже я подробно об этом напишу.
xenohunter
Кстати, на эту тему есть неплохая статья, правда, совсем «лёгкая».