

Ты мне — я тебе. Эта избитая фраза отлично описывает суть симбиоза, когда участники таких отношений, относящиеся к разным видам, получают какую-либо выгоду друг от друга. Примеров такой связи в природе можно встретить множество: анемоны и рыбы-клоуны, морские бычки и креветки, зебры и буйволовые скворцы и т.д. Одним из самых необычных симбиозов уверенно можно назвать связь между гавайским кальмаром и светящимися бактериями, обитающими в световом органе его мантии. Выгода для бактерий до ужаса банальна — пища, но вот кальмары получают от своих микроскопических напарников куда более удивительный дар — способность светиться. Удивительно и то, что молодняк кальмаров не обладает симбиотическими бактериями с рождения, а приобретает их в ходе фильтрации окружающей воды. Ученые из Калифорнийского университета (США) решили выяснить, что позволяет этим столь разным видам работать в паре. Какие механизмы лежат в основе симбиоза кальмара и бактерий, и как бактерии знают, где им поселиться? Пролить свет на эти вопросы нам поможет доклад ученых. Поехали.
Основа исследования

Главным героем данного труда, как мы уже знаем, является кальмар вида Euprymna scolopes. В отличие от своего колоссального собрата (Mesonychoteuthis hamiltoni), достигающего в длину 14 метров, этот вид не внушает ужаса своими габаритами — 3 см в длину весом 3 грамма. Обитает E. scolopes в водах рядом с Гавайскими островами и острова Мидуэй. Их можно встретить на глубинах от самой поверхности воды и до 240 метров. Продолжительность жизни также довольно мала — примерно 1 год (половая зрелость наступает уже через 2 месяца после вылупления).
В дневное время кальмары предпочитают прятаться, буквально с головой зарываясь в песок (специальная слизь, выделяемая кожей, помогает песчинкам прилипать и не скатываться).
Учитывая свои негабаритные габариты (простите за каламбур), врагов у гавайских кальмаров полно. Раз уж силу, скорость и внушительный вид эволюция тебе не подарила, необходимо проявить немного креативности и смекалки. E. scolopes живут в симбиозе с биолюминесцентными бактериями вида Aliivibrio fischeri (или же Vibrio fischeri). В начале жизненного пути бактерий в теле кальмаров нет, однако они попадают туда посредством фильтрации окружающей воды во время движения. В результате такого сожительства кальмары получают способность светиться в темноте, что очень полезно, учитывая их ночной образ жизни. Можно предположить, что это отвратительная маскировка, ибо кальмар должен привлекать массу внимания со стороны недоброжелателей, когда светится в ночи, словно новогодняя елка. Однако подобная тактика все же работает, поскольку хищники, находящиеся ниже кальмара, воспринимают его свечение как свет звезд на ночном небе.
Демонстрация биолюминесценции гавайского кальмара.
Факт того, что в теле кальмара обитают бактерии, сложно назвать уникальным, ибо практически у всех животных имеются такие попутчики. В теле человека обитает целая коллекция разных бактерий, вирусов и грибков. Естественно, часть микробиома несет вред, но большинство микроорганизмов участвуют в переваривании пищи, в процессах синтеза некоторых полезных веществ и т.д. Другими словами, это тоже является отличным примером симбиоза с теми же бактериями.
Изучение связи между организмом-носителем и микроорганизмами, его населяющими, уже проводилось. В предыдущих исследованиях ученые уделяли особое внимание роли, которую играет химическая коммуникация между видами.
К примеру, патоген растений Ralstonia solanacearum продуцирует гибридный липопептид нерибосомной пептид-синтетазы-поликетид-синтазы (NRPS-PKS), который вызывает развитие хламидоспор у нитчатых грибов, обитающих в почве, что способствует его сохранению в почвенной среде. А вот виды грибов Fusarium fujikuroi и Botrytis cinerea продуцируют бикаверин для подавления инвазии штаммов R. solanacearum, продуцирующих ралсоламицин.
В обоих случаях важнейшую роль для понимания взаимодействия между симбиотическими видами играли особые метаболиты*.
Метаболиты* — продукты метаболизма каких-либо соединений.По словам автором исследования, изучение бактерий V. fischeri позволяет расширить знания о биологически значимых соединениях, которые влияют на микробное поведение и передачу сигналов между микробом и носителем.
Вскоре после вылупления кальмар-хозяин получает симбионт из морской воды, и микроб начинает колонизировать покрытые эпителием крипты* светового органа.
Крипты* — узкие анатомические структуры, глубоко проникающие в более крупную структуру.В результате бактерии получают безопасное место жительства с трехразовым питанием, а кальмары — люминесценцию.
Чем так уникальна для ученых связь между кальмаром и бактериями? Тем что, система кальмар/бактерия является самой простой формой симбиоза, в которой имеется только два участника. Это позволяет анализировать специализированные метаболиты, участвующие в колонизации эпителиальных тканей. Кроме того, ткани кальмара практически прозрачны, что значительно облегчает процесс визуализации процесса колонизации тканей бактериями.
Уникальность еще и в том, что только V. fischeri (и только определенные штаммы V. fischeri) колонизируют световой орган E. scolopes. Частично это обусловлено обменом химическими сигналами между двумя партнерами, который приводит к активации симбиоза, включая высвобождение бактериальных продуктов пептидогликана, пептидогликанового мономера, трахеального цитотоксина (TCT), липополисахарида (LPS), в то время как хозяин выделяет оксид азота (NO) и хитин.
Основной контрольной точкой для этой специфичности является то, что производство бактериальной биопленки* необходимо для бактериальной агрегации и последующей колонизации. Бактериальные агрегаты были впервые обнаружены во время конфокальной визуализации колонизации кальмаров штаммом V. fischeri ES114. Было показано, что генетическая основа для образования агрегатов требует гибридной гистидинкиназы RscS, гибридной гистидинкиназы SypF, регулятора ответа SypG и 18-генный целевой полисахаридный (syp от symbiosis polysaccharide) локус. Штаммы, в которых отсутствуют какие-либо из вышеперечисленных регуляторов, не могут устойчиво колонизировать гавайских кальмаров.
Биопленка* — множество микроорганизмов, расположенных на какой-либо поверхности, клетки которых прикреплены друг к другу.Еще одно исследование показало наличие дополнительных регуляторов биопленки, включая BinK и HahK. BinK — сильный негативный регулятор* образования биопленок. У штаммов, лишенных BinK, наблюдается повышенное образование биопленок, тогда как сверхэкспрессия BinK приводит к снижению или отсутствию образования биопленок.
Негативные регуляторы* препятствуют транскрипции или трансляции. К примеру, cFLIP подавляет механизмы гибели клеток, приводящие к патологическим расстройствам.HahK передает сигнал NO от кальмара-носителя через датчик HnoX NO. В присутствии NO HnoX ингибирует активность HahK, способствуя образованию биопленки, что приводит к усиленному рассеянию симбиотической биопленки.
Ученые предполагают, что дополнительная продукция малых молекул (например, NO) может способствовать склонности V. fischeri вытеснять другие бактерии.
Характеристика биологических ролей положительного регулятора RscS и отрицательного регулятора BinK была облегчена с помощью анализов на основе культур, которые отражали симбиотические фенотипы. В частности, когда RscS сверхэкспрессируется (с использованием плазмидного или хромосомного аллеля rscS*), V. fischeri продуцирует большие агрегаты и вытесняет дикие типы у кальмаров.
Эти же штаммы образуют видимые биопленки на агаре, которые проявляются в виде морщинистых колоний. На другом конце спектра штаммы, у которых прерывается ген, кодирующий положительный регулятор RscS, неспособны образовывать биопленку.
Таким образом, используя набор генетически измененных производных V. fischeri ES114, удалось изучить спектр фенотипов биопленок в культуре, которые имеют отношение к симбиотическому поведению хозяина.
В ходе исследования были выявлены три основные изогенные (генетически идентичные) штаммы:
- I — штамм, который не образует симбиотическую биопленку в культуре или у животных («нет биопленки», т.е. ES114 rscS* rscS::Tnerm). Этот штамм содержит прерывание в rscS, которое блокирует передачу сигналов биопленки.
- II — штамм, который образует симбиотическую биопленку у животного, но не в культуре («дикий тип» (WT от wild type); т.е. неизмененный ES114).
- III — штамм, который образует симбиотическую биопленку в культуре и образует усиленные агрегаты у животных из-за сверхэкспрессии положительного регулятора RscS и отсутствия отрицательного регулятора BinK («есть биопленка», т.е. ES114 rscS* DbinK).
Результаты исследования
В ходе первого этапа исследования была использована масс-спектрометрия с визуализацией на агаре (IMS от agar-based imaging mass spectrometry), позволившая получить скрининг отношения массы к заряду (m/z) у штаммов V. fischeri WT с биопленкой и без биопленки для идентификации ионов, которые различаются между колониями трех штаммов на агаре.
Модовый анализ в диапазоне малых молекул (от 100 до 1000 Да) позволил получить диаграмму масс, которые были значительно более многочисленными в штамме III, чем в двух других образцах. Один из сигналов (m/z 257) был обнаружен с высокой интенсивностью в штамме III и c низкой интенсивностью в штамме WT и в штамме I (изображение №1).
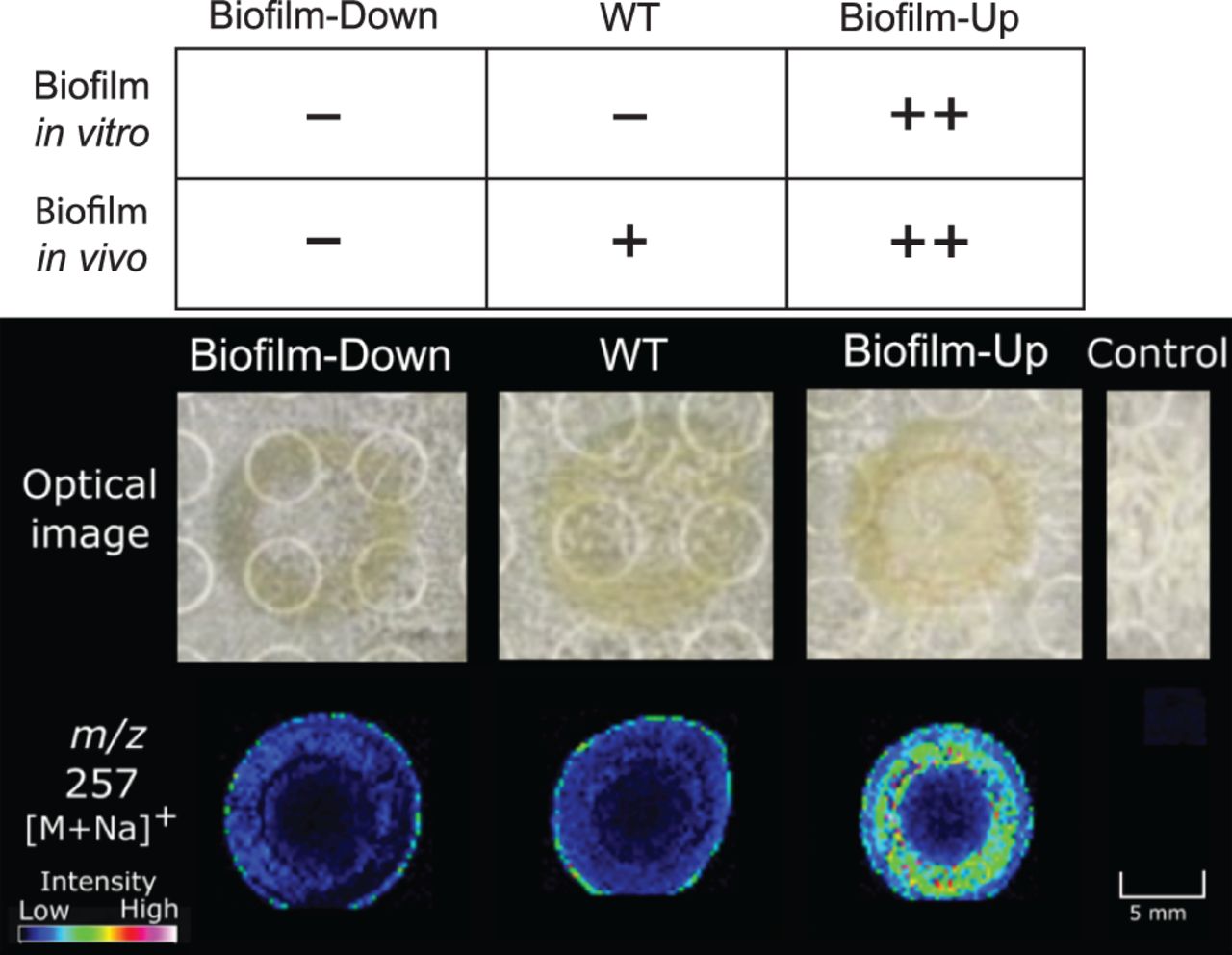
Изображение №1
Далее была выполнена проверка этого соединения с помощью кальция, который может стимулировать образование биопленок V. fischeri у штаммов, лишенных BinK без необходимости сверхэкспрессии rscS*.
В ES114 наблюдалось повышенное содержание соединения в кальциевой среде после индукции биопленки (т.е. в штамме, лишенном BinK), что подтверждает использование этой методики анализа. Также было обнаружено повышенное содержание соединения в индуцированных биопленками штаммах MB11B1, MB15A4 и SR5.
Таким образом, эти результаты показывают, что это соединение обнаруживается в нескольких моделях биопленок и может быть обнаружено в штаммах, представляющих три основные филогенетические группы симбиотических V. fischeri.
Массы, идентифицированные с помощью IMS, были занесены в список путем оценки их статистической значимости.
Поскольку ионизационная способность изменяется при различных модальностях, данные жидкостной хроматографии и тандемной масс-спектрометрии неочищенных экстрактов V. fischeri из каждого мутанта были оценены с помощью международной базы спектрометрии GNPS (Global Natural Products Social). Это позволило исследовать сигналы, обнаруженные в IMS, а также изучить соединения, которые могут ионизироваться только электрораспылением (ESI от electrospray ionization).
Спектральное соответствие экстракта биопленки V. fischeri было обнаружено для небольшой молекулы цикло (гистидил-пролина) (cyclo(His-Pro) или cHP) с молекулярной массой 234 г/моль.
Данные анализа показали, что m/z 257, вероятно, является натриевым аддуктом* cyclo(His-Pro) (C11H14N4O2), а протонированной* молекулой была m/z 235.
Аддукт* — продукт прямого присоединения молекул друг к другу в химической реакции.
Протонирование* — перенос протона с кислоты на протофильную частицу.Cyclo(His-Pro) является членом структурного класса дикетопиперазинов* (DKP от diketopiperazine), которые образуются в результате циклизации* двух аминокислот и являются преобладающими натуральными продуктами, которые играют различные роли в микробных взаимоотношениях.
Дикетопиперазин* — класс органических соединений, родственных пиперазину, но с двумя амидными связями.
Циклизация* — процесс образования циклических соединений, т.е. химических соединений, в которых присутствует три или более связанных атомов, образующие кольцо.

Изображение №2
На графике выше показан результат прямой инфузии, выполненной для обнаружения и подтверждения моделей фрагментации m/z 235 из экстракта биопленки V. fischeri и коммерческого cyclo(L-His-L -Pro). Тут видно практически идеальное совпадение между моделями фрагментации обоих образцов, что дополнительно подтверждается с помощью масс-спектрометрии высокого разрешения с ионизацией электрораспылением (HRESIMS от high-resolution electrospray ionization mass spectrometry).
Чтобы выяснить стереохимическую конфигурацию DKP, было синтезировано все четыре возможных стереоизомера cyclo(His-Pro): cyclo(L-His-L-Pro) (cHP -1); cyclo(L-His-D-Pro) (cHP-2); cyclo(D-His-L-Pro) (cHP-3) и cyclo(D-His-D-Pro) (cHP-4). Четыре стереоизомера были синтезированы для химического сравнения с выделенным соединением из экстракта биопленки V. fischeri (3А).

Изображение №3
Для согласования времени удерживания DKP в экстракте V. fischeri со всеми четырьмя синтезированными стереоизомерами использовалась хиральная хроматография* (3B).
Хиральная хроматография* – метод разделения оптических изомеров на хиральных неподвижных фазах (ХНФ).Анализ показал, что пик стереоизомера cHP-3 также обнаруживался в экстракте биопленки V. fischeri. Сопоставление обоих пиков приводило к их практически полному слиянию, что подтверждало идентичность их структуры и конфигурации (3C). Другие стереоизомеры были исключены как продуцируемые в экстракте биопленки, так как время их удерживания не соответствовало существующему пику в экстракте биопленки либо пик совместной инъекции не сливался с пиком экстракта. Количественная оценка молекулы в экстрактах биопленок, продуцируемых мутантами, также показала, что мутант с биопленкой продуцировал большее количество cHP.
Дабы лучше понять роль cHP-3 в симбиозе бактерий и кальмара-носителя, необходимо было проанализировать продукцию cHP-3 в реальном времени.
Сначала ученые проверили, генерируется ли cHP-3 в световых органах недавно вылупившихся особях E. scolopes. В ходе данного тестирования было проверено три условия: отсутствие V. fischeri (апосимбиотик); V. fischeri WT (дикий тип) и V. fischeri WT DbinK (аналог штамма I, но вместо генетического индуцирования образования биопленки с аллеля rscS*, данный штамм зависит от индукции нативных сигналов в кальмаре-носителе).
Вылупившиеся E. scolopes инокулировали каждым образцом в течение 3 часов, промывали и позволяли продолжить колонизацию в течение 48 часов. Через 48 часов m/z 235 в контрольной группе (без V. fischeri) вообще не был обнаружен, в группе WT — были слабые сигналы, но вот в третьей группе (с V. fischeri WT DbinK) продукция была самой высокой (изображение №4).

Изображение №4
Эти результаты являются доказательством того, что молекула продуцируется штаммом WT и что нокаут binK приводит к увеличению продукции иона.
Проведение подобного теста в пробирке и в живом организме показало, что биопленки генетически индуцируются аллелем rscS* в пробирке, но в живом организме этот аллель отсутствует. Причина этого различия заключается в том, что активированный аллель RscS используется для имитации поведения в культуре, которое стимулируется условиями особи-носителя. Следовательно, RscS не является необходимостью для активации биопленки, т.е. для формирования симбиоза.
Далее необходимо было понять роль обнаруженной молекулы в люминесценции.
В работе коллективных бактерий важную роль играет чувствительность кворума (QS от quorum sensing) — активность бактериальных клеток, участвующих в групповом поведении, чему способствует выработка QS молекул отдельными клетками до тех пор, пока не будет достигнута необходимая концентрация для активации определенного путь (т.е. поведения). В случае V. fischeri активируется путь LuxR и возникает биолюминесцентная активность.
Учитывая связь между потенциалом производства биопленок и появлением cHP-3 в колонизированном носителе, необходимо было установить роль именно cHP-3 в биолюминесценции V. fischeri. Увеличение люминесценции наблюдалось при низкой плотности клеток у штамма WT (ES114) (изображение №5).

Изображение №5
Поскольку ES114 имеет низкую люминесценцию по сравнению с другими изолятами V. fischeri, действие cHP-3 было протестировано на другом штамме (EM17), который является гораздо более ярким изолятом. Этот штамм также показал усиление люминесценции при добавлении экзогенного cHP-3, и этот эффект был гораздо более выраженным, чем у ES114. Наибольшее влияние на люминесценцию у обоих штаммов наблюдалось при концентрациях cHP-3 100 и 250 мкМ.
Однако на этом роль cHP-3 в симбиозе между бактериями и кальмаром не ограничивается. Наблюдаемое увеличение продукции cHP-3 в мутанте с образованием биопленки может свидетельствовать о том, что cHP-3 играет решающую роль в развитии биопленки, либо как конечная цель, либо как предварительный шаг для последующего процесса, или даже как QS молекула.
Для более подробного ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
В данном труде ученые исследовали симбиотическую связь между кальмарами и светящимися бактериями, которые делятся с носителем своей биолюминесценцией в обмен на питательные вещества.
В рамках анализа симбиоза между, грубо говоря, крупными организмами редко возникают вопросы касательного того, что является стимулом к началу симбиоза. Однако, когда одним из симбионтов является бактерией, мотив партнерства ясен, но вот понять механизм, стоящий за ее поведением, куда сложнее.
Данный труд показал, что основную роль в формировании симбиоза между кальмарами и бактериями играет маленькая молекула цикло(D-гистидил-L-пролин) или просто cHP-3, относящаяся к дикетопиперазинам (DKP).
Эта молекула участвует в формировании биопленки и в люминесцентном поведении бактерий, которые подчиняются законам коллективизма. Другими словами, отсутствие данной молекулы привело бы к тому, что бактерии попросту не светились бы, а может и не поселялись бы в кальмарах. Ученые уверены, что данная молекула вырабатывается в течение первых часов симбиотической связи.
По своей сути, cHP-3 является химическим сигналом, специфичным для этого конкретного симбиоза, но пока не был установлен источник этих молекул. В дальнейшем ученые намерены провести дополнительные исследования, чтобы выявить гены, отвечающие за производство cHP-3.
Симбиоз это взаимовыгодное сосуществование, но далеко не всегда один из участников готов осознанно вступать в такие отношения. В случае с кальмарами и бактериями причиной является факт того, что у бактерий попросту нет сознания, нет глаз, носа или других классических органов чувств. Следовательно, для привлечения такого необычного, но жизненно важного напарника кальмары используют химические сигналы в виде молекул.
Столь удивительное партнерство между столь удивительными существами лишний раз показывает, как мало мы знаем об окружающем нас мире, особенно об обитателях морских глубин.
Пятничный офф-топ:
В мире дикой природы много примеров симбиоза. Одним из самых необычных является партнерство между муравьями Pseudomyrmex ferruginea и растением Vachellia cornigera, которое они оберегают, как зеницу ока.
В мире дикой природы много примеров симбиоза. Одним из самых необычных является партнерство между муравьями Pseudomyrmex ferruginea и растением Vachellia cornigera, которое они оберегают, как зеницу ока.
Офф-топ 2.0:
Симбиоз, который продолжается далеко не первое поколение, приводит к определенным эволюционным изменениям. К примеру, лапки буйволовых скворцов устроены так, чтобы цепляться к крупным млекопитающим под любым углом и не сваливаться при движении.
Симбиоз, который продолжается далеко не первое поколение, приводит к определенным эволюционным изменениям. К примеру, лапки буйволовых скворцов устроены так, чтобы цепляться к крупным млекопитающим под любым углом и не сваливаться при движении.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
drWhy
Любопытно и познавательно, как и всегда. Спасибо.