
 В предыдущей части мы показали, что в клеточном автомате могут возникать волны, имеющие специфический внутренний узор. Такие волны могут запускаться из любого места клеточного автомата и распространяться по всему пространству клеток автомата, перенося информацию. Соблазнительно предположить, что реальный мозг может использовать схожие принципы. Чтобы понять возможность аналогии, немного разберемся с тем, как работают нейроны реального мозга.
В предыдущей части мы показали, что в клеточном автомате могут возникать волны, имеющие специфический внутренний узор. Такие волны могут запускаться из любого места клеточного автомата и распространяться по всему пространству клеток автомата, перенося информацию. Соблазнительно предположить, что реальный мозг может использовать схожие принципы. Чтобы понять возможность аналогии, немного разберемся с тем, как работают нейроны реального мозга.Головной мозг состоит из серого и белого вещества. Серое вещество – это мозговые структуры, состоящие из нейронов и глиальных клеток. Белое вещество – это аксоны нейронов, они же нервные волокна. Эти волокна образуют связи одних структур мозга с другими.
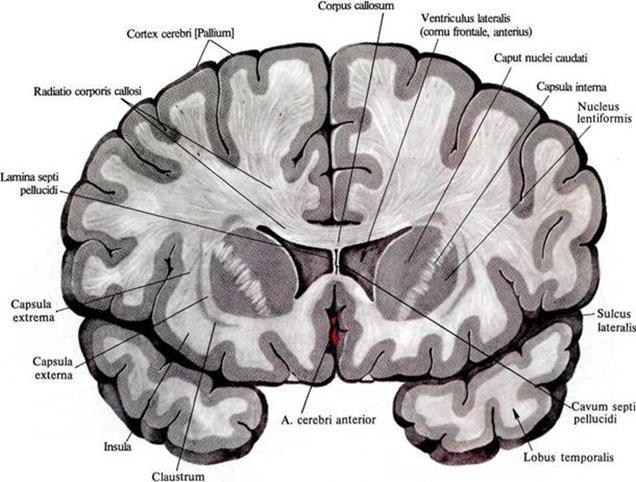
Распределение белого и серого вещества на фронтальном срезе мозга
Структуры, которые находятся ближе к центру мозга, принято относить к древнему мозгу. Древний мозг роднит нас с животными и реализует механизмы, отточенные эволюцией и более-менее общие для многих живых существ. Основной объем серого вещества человека приходится на кору. Кора – это слой серого вещества толщиной от 1.3 до 4.5 мм, составляющий наружную поверхность мозга. Есть множество доводов в пользу того, что кора, в отличие от древнего мозга, реализует не генетически заложенные алгоритмы, а способна к обучению и самоорганизации.
Основные клетки мозга – это нейроны и глиальные клетки. Похоже, что и те, и другие играют существенную роль в информационных процессах. Для упрощения повествования, пока, мы будем говорить только о нейронах. Разговор же о глиальных клетках отложим на некоторое время.
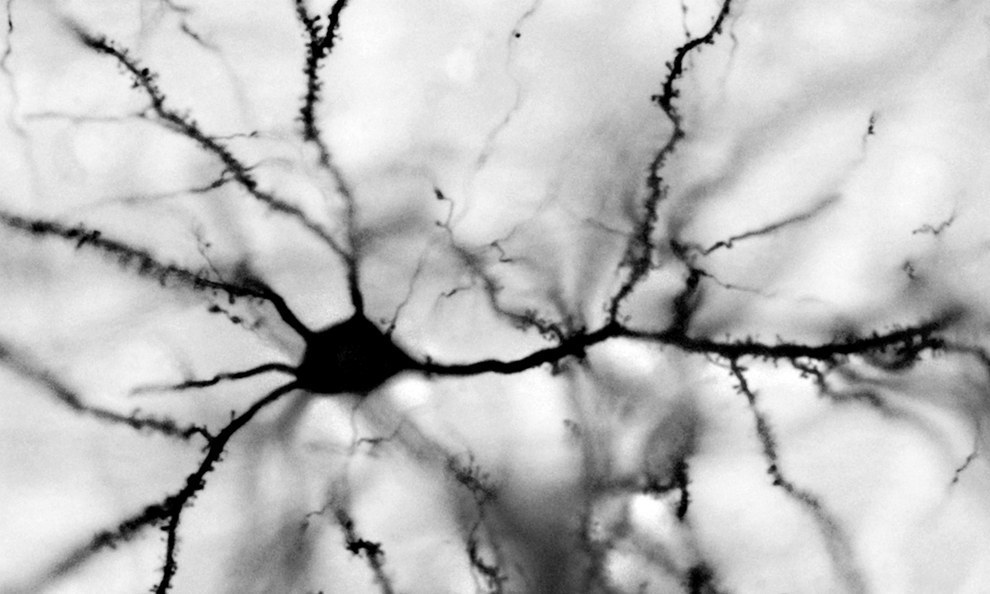
Нейроны бывают разных типов. Самые массовые нейроны коры — это пирамидальные нейроны. На их долю в коре приходится 75% от всех нейронов. На рисунке ниже показаны именно они.
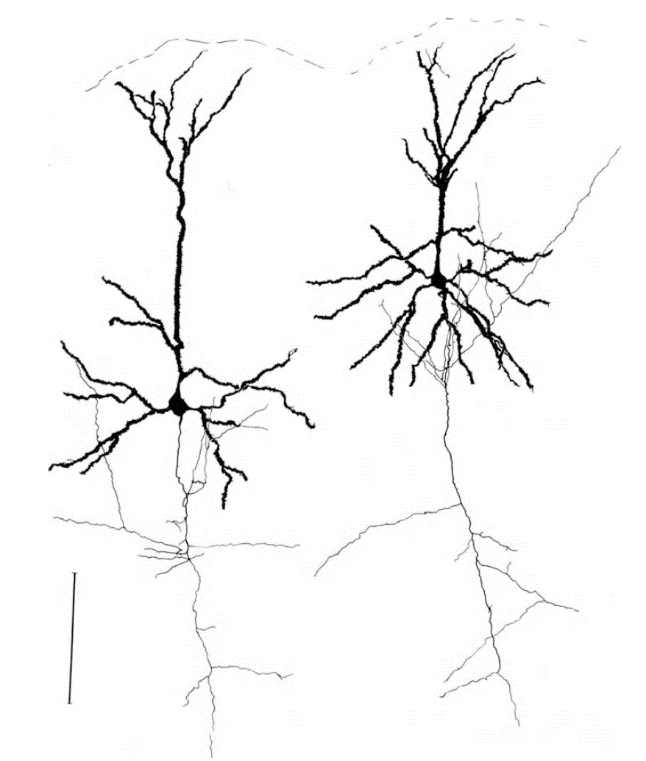
Структура пирамидального нейрона, черный – дендрит, серый – аксон, линейка – 0.1 мм (Braitenberg, 1978)
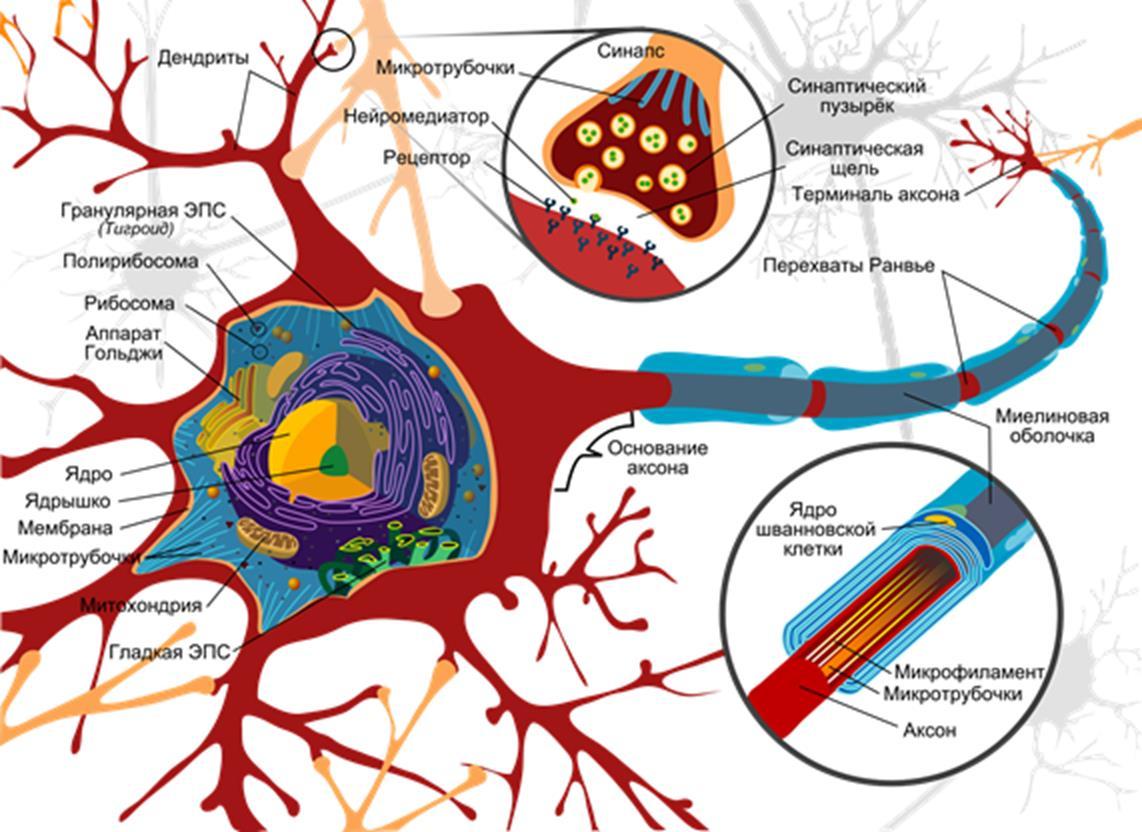
У большинства нейронов есть тело, дендритное дерево и аксон. И аксон, и дендрит сильно ветвятся и образуют запутанную структуру со множеством переплетений с дендритами и аксонами других нейронов. Составить общее представление о сложности и запутанности переплетений аксонов и дендритов можно, например, по видео.
Общую конфигурацию нейрона хорошо представить по классической картинке из википедии.
Тела нейронов, их дендриты и аксоны, окружающие глиальные клетки все они тесно упакованы между собой, оставляя свободными только узкие щели. Эти щели заполнены сложным раствором существенную часть которого составляют электролиты (в основном, ионы калия, кальция, натрия и хлора). Плотность упаковки можно увидеть и оценить на реконструкции небольшого объема коры, приведенной ниже.

(connectomethebook.com)
Поверхность нейрона называется мембраной. Задача мембраны оградить внутреннюю среду нейрона от внешней. При этом в мембрану встроено огромное количество белков. Некоторые из них пронзают мембрану насквозь и контактируют, как с внешней, так и внутренней средой нейрона. Такие белки называют трансмембранными (рисунок ниже).
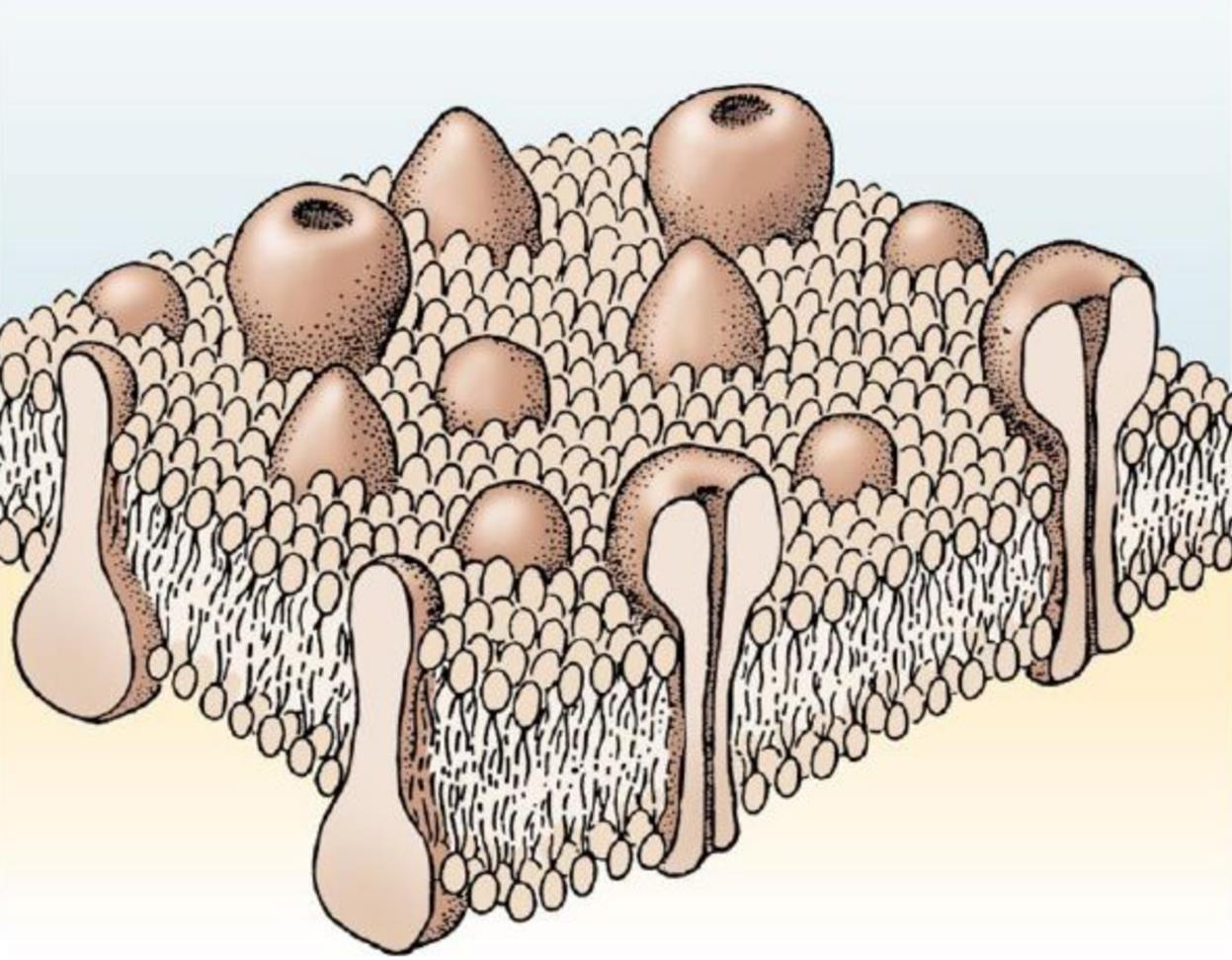
Трансмембранные белки
Трансмембранные белки выполняют разные функции. Если белки занимаются транспортировкой ионов в клетку или из клетки и делают это постоянно, то это ионные насосы. Чтобы осуществлять транспорт ионов белки создают ионные каналы. Ионные каналы могут иметь внешнее управление, то есть открываться и закрываться при определенных сигналах. Если канал управляется мембранным потенциалом, то говорят о потенциал-зависимых ионных каналах.
Если белок реагирует на какое-либо вещество снаружи нейрона и передает эту реакцию каким-либо способом внутрь нейрона, то такие белки называют рецепторами. Вещество, которые воздействуют на конкретный рецептор называют его лигандом. Если рецептор имеет ионный канал, который открывается под воздействием лиганда, то такой рецептор называется ионотропным. Если у рецептора нет ионного канала и он воздействует на состояние нейрона окольным путем, то это метаботропный рецептор.
Рецепторы и другие белки не сосредоточены где-то в одном месте, а распределены по всей поверхности нейрона. Средний нейрон коры имеет порядка 10 000 синапсов, распределенных по его дендриту и телу. На каждый синапс приходится несколько сотен рецепторов.
В состоянии покоя между внутренней и внешней средой нейрона существует разность потенциалов – мембранный потенциал, составляющий около 70 милливольт. Он образуется за счет работы белковых молекул, работающих как ионные насосы. В зависимости от своего типа ионные насосы меняют соотношение определенных ионов снаружи и внутри клетки. Насосы первого типа меняют соотношение ионов калия и натрия, второго типа – выводят из клетки ионы кальция, третьего типа – транспортируют наружу протоны. В результате мембрана приобретает поляризацию, при которой отрицательный заряд накапливается внутри клетки, а положительный снаружи.
Места контактов аксонов с дендритами или телами нейронов называются синапсами. Основной тип синапсов – это химический синапс.
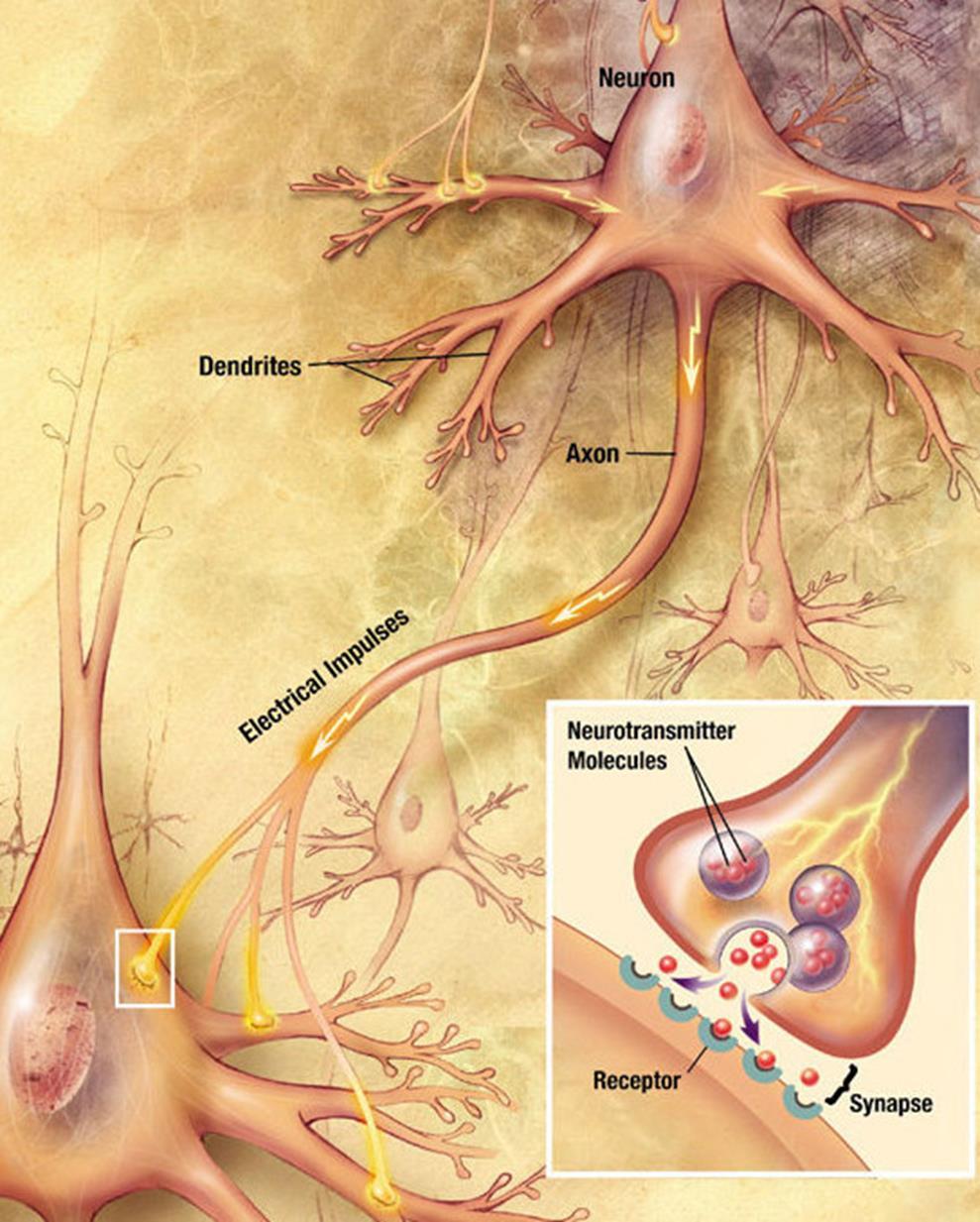
Когда нервный импульс по аксону поступает в синапс, он высвобождает из специальных пузырьков молекулы нейромедиаторов, характерные для этого синапса. На мембране нейрона, получающего сигнал, есть белковые молекулы – рецепторы. Рецепторы взаимодействуют с нейромедиаторами. Рецепторы, расположенные в синаптической щели, являются ионотропными, то есть они же являются ионными каналами, способными перемещать ионы. Нейромедиаторы так воздействуют на рецепторы, что их ионные каналы открываются. Соответственно, мембрана либо деполяризуется, либо гиперполяризуется – в зависимости от того, какие каналы затронуты и, соответственно, какого типа этот синапс. В возбуждающих синапсах открываются каналы, преимущественно пропускающие катионы внутрь клетки, — мембрана деполяризуется. В тормозных синапсах открываются каналы, выводящие катионы из клетки, что приводит к гиперполяризации мембраны.
Поляризация мембраны нейрона выглядит, как скопление ионов в относительной близости от мембраны (рисунок ниже).
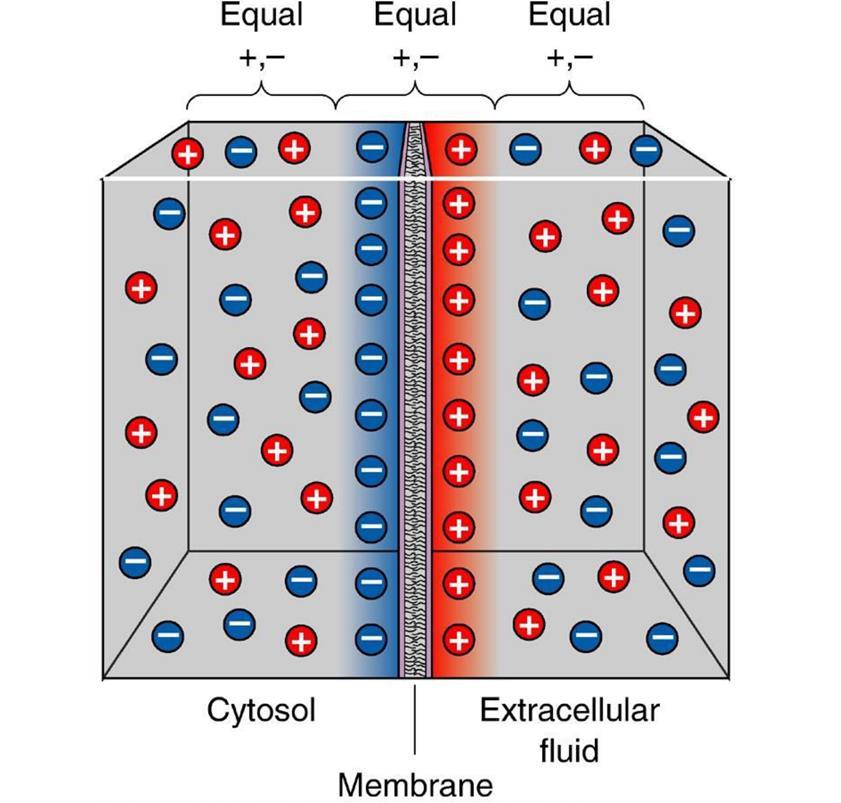
Когда открываются ионные каналы рецепторов и начинается обмен ионами с внешней средой, то только в том месте поверхности нейрона где расположены рецепторы и происходит изменение поляризации. Маленький участок мембраны оказывается заряжен иначе чем все его окружение.
Если сработали возбуждающие рецепторы, то участок в соответствующем месте окажется деполяризован, то есть его потенциал будет выше чем в среднем по мембране нейрона. Если эта деполяризация достигнет критического значения, то возникнет спайк, который начнет распространяться по мембране.
За возникновение и распространение спайка отвечают потенциал-зависимые ионные каналы. Они управляются не нейромедиаторами, а величиной мембранного потенциала. Например, для аксона их работа выглядит следующим образом.
Когда потенциал поднимается до критического значения открываются натриевые каналы, которые начинают гнать положительно заряженные ионы натрия внутрь клетки. Соответственно, потенциал в этом месте лавинообразно возрастает. Но в определенный момент включаются калиевые потенциал-зависимые каналы. Они начинают выводить положительно заряженные ионы калия из клетки наружу, понижая тем самым мембранный потенциал. В результате происходит кратковременный локальный всплеск потенциала. Затем наступает рефрактерный период, когда это место оказывается нечувствительно к изменению потенциала. Но сильный всплеск в одном месте приводит к менее сильному подъему потенциала в соседних с ним местах. Там происходит превышение порогового значения и рождается свой всплеск. В результате потенциал действия или иначе спайк распространяется по всей длине аксона.
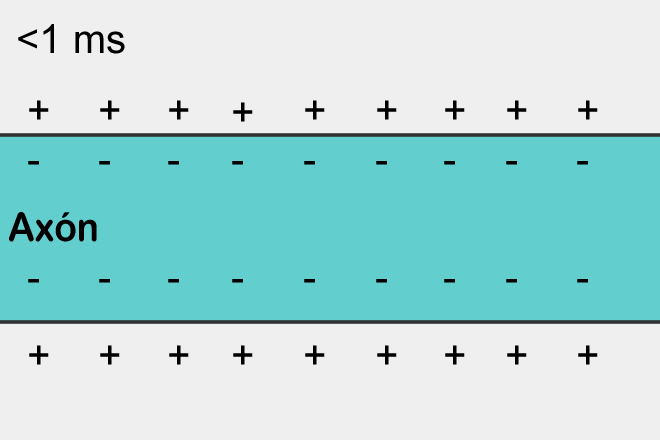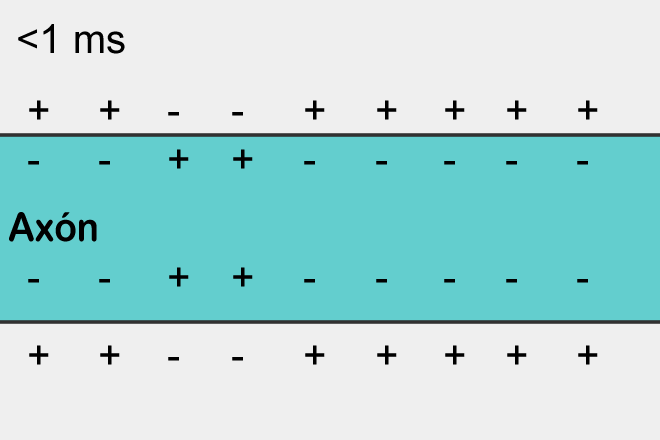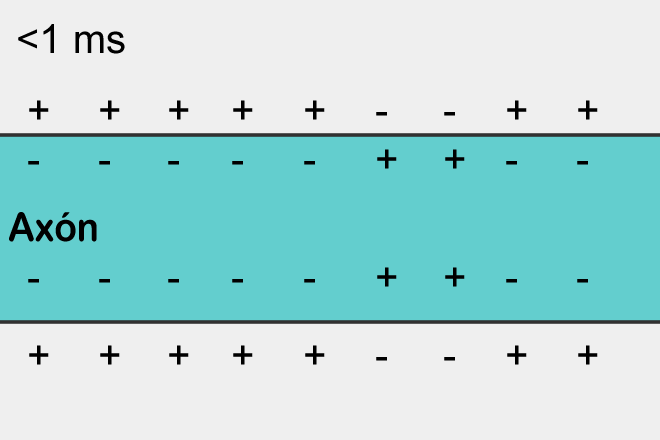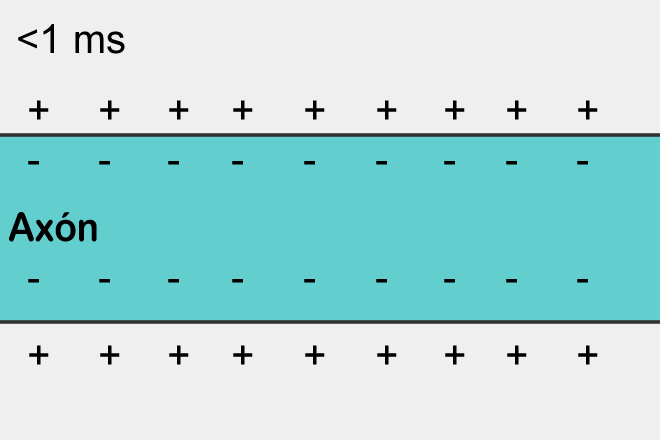
Распространение спайка – самовоспроизводящийся процесс. Спайк, возникнув в одном месте, заставляет соседние с ним места сгенерировать свой спайк и так далее. Это, кстати, напоминает простейший клеточный автомат, наподобие того, что мы описывали в предыдущей части. Возникнув в одном месте спайк распространяется во все стороны от этого места. Но если спайк не возник в этом месте, а пришел со стороны, то за счет того, что есть рефракторный период, он может распространить только туда, где его еще не было.
В аксонах покрытых миелиновой оболочкой потенциал действия распространяется несколько иначе. Миелиновая оболочка не дает распространяться спайку, но, зато, хорошо изолирует нервное волокно. В результате, внутри изолированной части происходит передача электрического сигнала, как по кабелю. Затем в неизолированном участке, перехвате, происходит генерация нового спайка. За счет таких «прыжков» скорость передачи нервного импульса в толстых, покрытых миелином, аксонах оказывается много выше, чем в нервных волокнах без такой оболочки.
В дендритах также есть потенциал-зависимые ионные каналы и по ним, как и по аксону может распространяться потенциал действия. Аксонные спайки имеют амплитуду порядка 100 мВ, амплитуда дендритных спайков несколько ниже. Аксонные спайки возникают на теле нейрона в дендритном холмике. Оттуда они распространяются далее по аксону. Возбуждение в аксонном холмике может распространяться и на дендрит, в этом случае возникают дендритные спайки, которые являются сигналом обратного распространения по отношению к аксонному потенциалу действия.
Дендритные спайки могут возникать и непосредственно в дендрите. Для этого требуется, чтобы в течении короткого временного интервала (порядка 3-10 мс) на небольшом участке дендрита случилось достаточно много синаптических возбуждений. Например, если длина участка 100 мкм и временной интервал 3 мс, то потребуется, чтобы сработало около 50 синапсов, чтобы появился дендритный спайк. Стоит учесть, что на таком участке располагается порядка 200 синапсов. Добиться синхронной активности четверти всех синапсов можно при искусственном возбуждении «в пробирке». Трудно сказать возможно ли такое в живой ткани.
Распространение спайков не единственный механизм передачи информации, свойственный дендриту. Показано, что дендриты обладают кабельными свойствами. Ветку дендрита можно сопоставить с кабелем, у которого есть внутреннее сопротивление, сопротивление утечки и поверхностная емкость. Хотя сопротивление дендрита и очень велико, а утечки существенны, тем не менее, токи, которые возникают от возбуждающих постсинаптических потенциалов могут оказывать существенное влияние на общее состояние нейрона. Можно предположить, что особенно роль этих токов велика на коротких расстояниях, например, в пределах одной веточки дендритного дерева.
И аксон, и дендритные ветки представляют собой тонкие трубочки. Распространение спайка по ним – это перемещение кольцевой области деполяризации. Но спайки – это достаточно энергетически затратные явления. Кроме них есть более слабые, но зато и более массовые сигналы. Нейробиологи, порою, говорят, что нейроны, в основном, не кричат друг другу (подразумевая спайки), а шепчут.
Вернемся к химическому синапсу. Нервный импульс, распространяясь по аксону, доходит до многочисленных аксонных терминалей. Большая часть терминалей образует контакты с дендритами. Это и есть химические синапсы. Дойдя до терминали, спайк вызывает массовый выброс нейромедиаторов в синаптическую щель. Нейромедиаторы упакованы в специальные пузырьки (везикулы). В одной везикуле содержится несколько тысяч молекул.
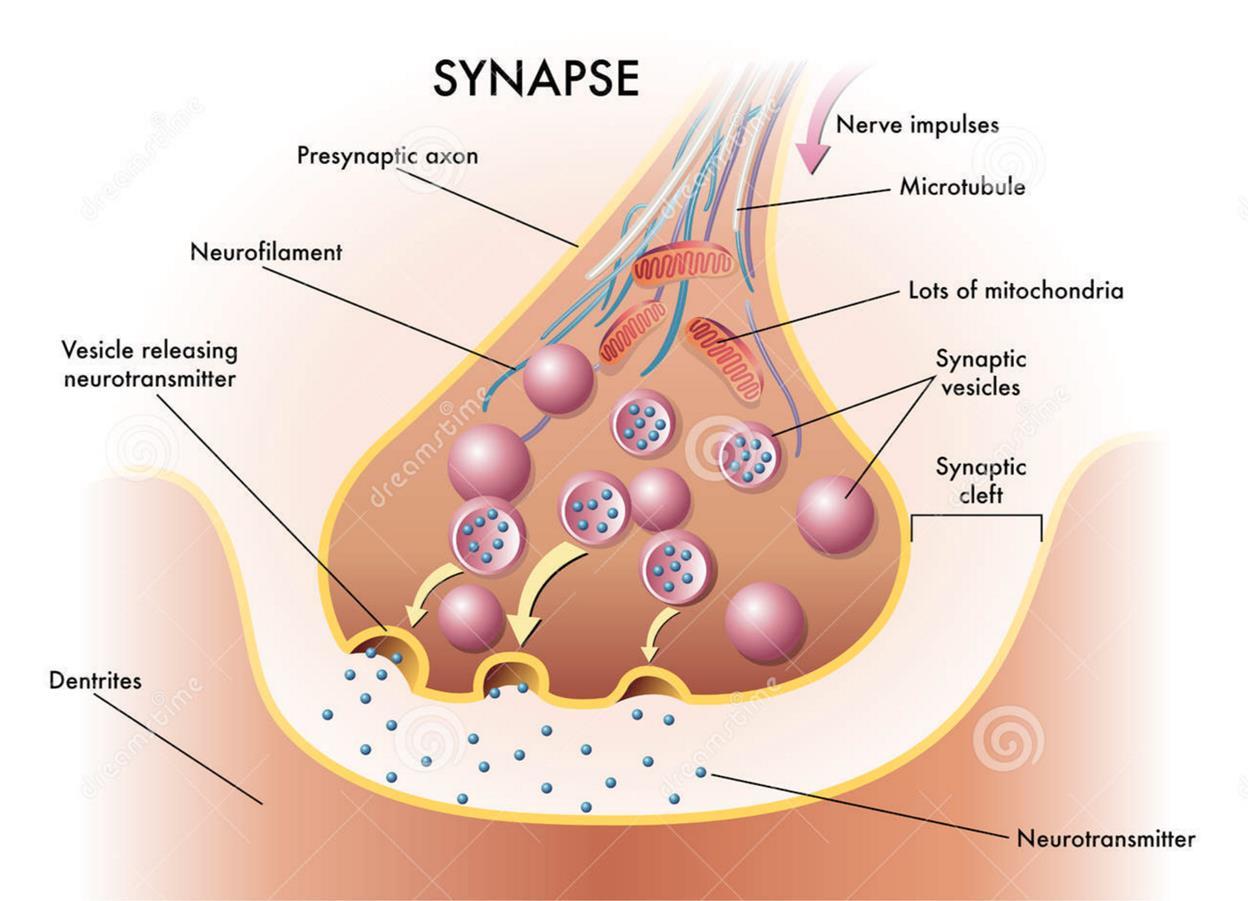
Приход спайка вызывает массовый выброс нейромедиаторов, состоящий из десятка пузырьков. Дозу нейромедиаторов, содержащуюся в одном синаптическом пузырьке принято называть квантом нейромедиатора.
Кроме массового выброса нейромедиаторов, происходящего в момент прихода аксонного спайка, есть еще, так называемое, квантовое испускание, когда выделяется только одна везикула с нейромедиаторами. Причем, квантовая активность может быть не связана с вызванной активностью нейронов к которым относится синапс и происходить независимо от нее.
Измерения, проделанные рядом с синапсами, показывают, что на мембране рядом с каждым синапсом время от времени фиксируются возбуждающие постсинаптические потенциалы, имеющие амплитуду порядка 1 мВ или кратную. Считается, что такие миниатюрные постсинаптические потенциалы связаны именно с квантовым испусканием нейромедиаторов.
Когда нейромедиаторы выбрасываются в синаптическую щель, то часть медиаторов попадает за пределы синаптической щели и растекается по пространству, образованному нейронами и окружающими их глиальными клетками. Это явление называется спиловером (spillover). Кроме того, медиаторы испускаются несинаптическими терминалями аксонов и глиальными клетками (рисунок ниже).

Источники медиаторов за пределами синаптической щели (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
Когда на дендрите нейрона что-то происходит это сопровождается выделением нейромедиаторов. Внутри синапса нейромедиаторы влияют на ионотропные рецепторы и как следствие на локальное изменение мембранного потенциала дендрита. Когда же нейромедиаторы попадают за пределы синапса они начинают влиять на все, что находится в непосредственной близости. При этом не важно есть ли между этими элементами непосредственный контакт. Это можно сравнить с толпой людей. Люди в толпе могут разбиться на пары и вести между собой беседы, но слышать эти беседы будут не только сами собеседники, но их ближайшие соседи.
Еще про синапсы надо сказать, что в их синаптических пузырьках хранится не один нейромедиатор, а определенный коктейль. Как правило, это смесь из одного основного нейромедиатора и нескольких дополнительных нейропептидов, которые называют нейромодуляторами. Таким образом, спиловер выбрасывает из синапса наружу целый набор сигнальный веществ. Разные нейроны одного типа могут иметь общий основной нейромедиатор, но при этом различаться составом дополнительных.
Большинство синапсов, где-то три четверти их числа, располагаются на небольших отростках, называемых шипиками. Шипики отодвигают синапс от дендрита и создают в пространстве такое распределение синапсов, что синапсы от разных дендритов оказываются перемешаны между собой.
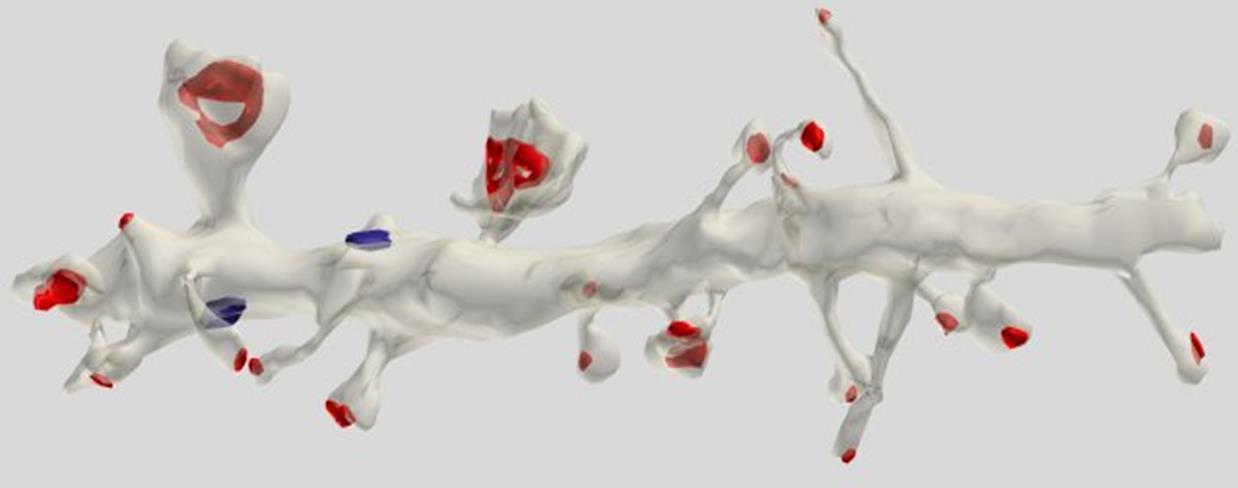
Реконструкция участка дендрита пирамидальной клетки. Красным отмечены синапсы на шипиках, синим – на дендритном стволе (Dr. Kristen M. Harris)
Если взять участок дендрита длиною 5 мкм (рисунок ниже), то на нем окажется порядка десяти синапсов. Но дендритные ветки одних нейронов тесно переплетаются с ветками других нейронов. Все они проходят друг от друга в самой непосредственной близости. В цилиндрический объем высотою 5 мкм и диаметром тоже 5 мкм попадает около 100 синапсов. То есть в 10 раз больше того количества, что непосредственно располагается на самой дендритной ветке.
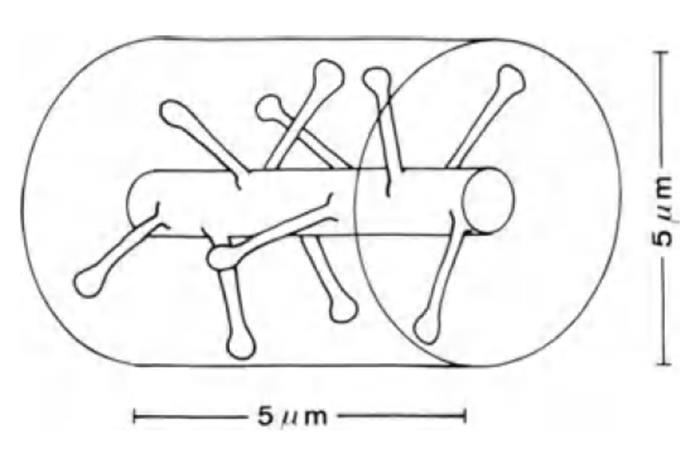
Участок дендрита (Braitenberg V., Schuz A., 1998)
В результате, синапсы образуют систему распределенных случайным образом источников нейромедиаторов для внесинаптической среды. Любая активность в синапсах вызывает появление нейромедиаторов в окружающем их пространстве. Если где-то одновременно становятся активны несколько соседних синапсов, то в таком месте возникает коктейль из тех медиаторов, что выделились из этих синапсов.
Если взять любое место, то поблизости друг от друга в радиусе полутора микрометров окажется около 10 синапсов. Большинство из них будет принадлежать разным дендритам. Если пронаблюдать за тем, какие сочетания нейромедиаторов будут появляться в этом месте, то окажется, что по составу «коктейля» можно каждый раз достаточно точно сказать какие именно синапсы были активны.
Можно привести такой пример. Представьте, что в округе есть 10 баров. Всего существует сотня сортов пива. В каждом разливают только по 3 сорта пива. Эти сорта в каждом баре бармен выбрал однажды случайным образом и теперь разливает только их. Вы обходите несколько баров, выпиваете в каждом по три пива разного сорта и забираете с собой картонные подставки под стаканы с названием выпитого пива. В итоге, по сочетанию картонок, почти всегда, вашей жене удастся определить в каких барах вы побывали.
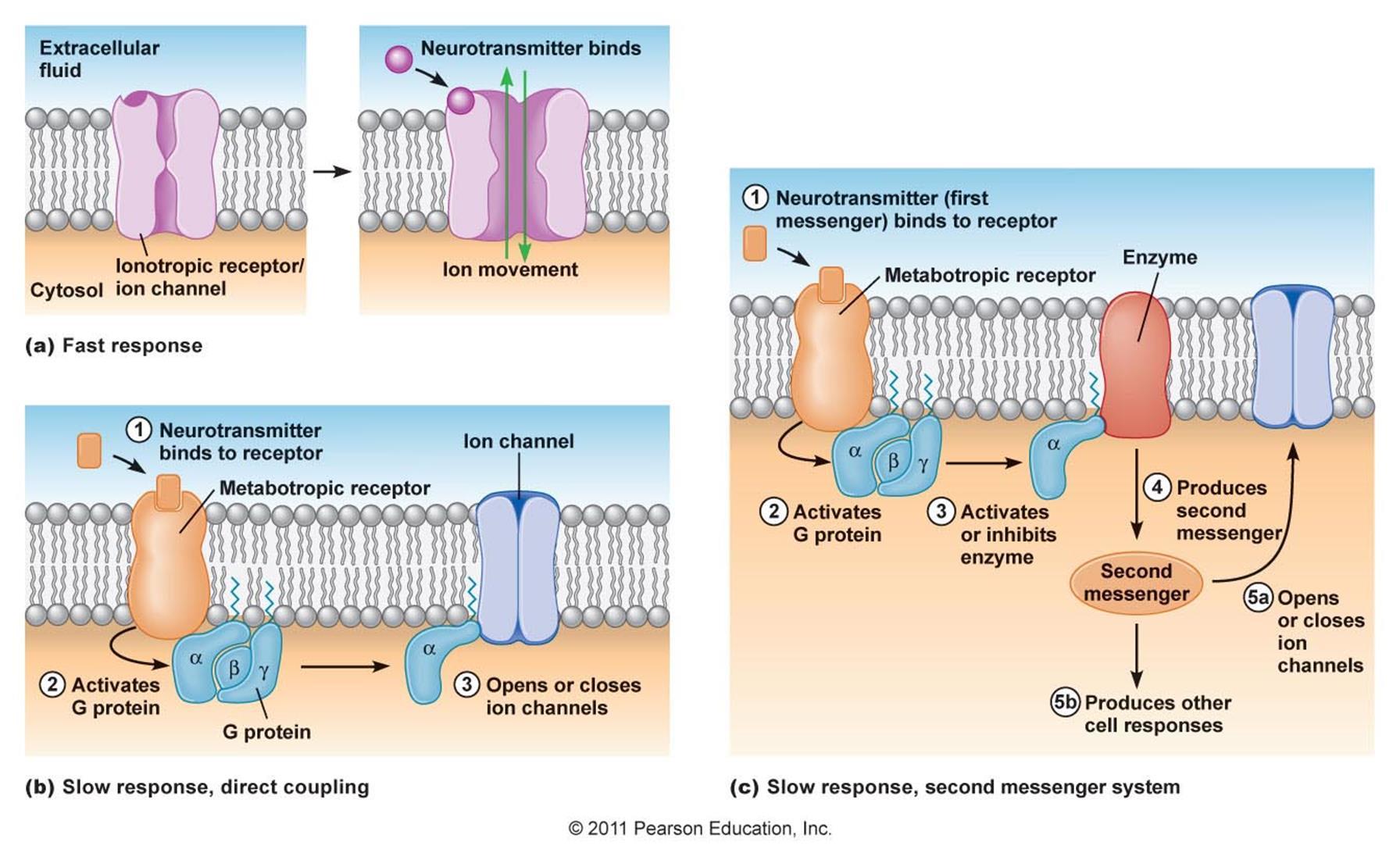
Нейромедиаторы, оказавшиеся вне синапсов имеют свой специфический механизм, позволяющий влиять на работу нейронов. На поверхности дендрита и тела нейрона в больших количествах располагаются метаботропные рецепторы. Эти рецепторы не имеют ионных каналов и не могут напрямую влиять на мембранный потенциал нейрона. С внутренней стороны мембраны эти рецепторы связаны с так называемым G-белком. За это их часто так и называют — рецепторы, сопряженные с G-белком (G-protein-coupled receptors, GPCRs). Когда метаботропные рецепторы активируются своим лигандом они отпускают G-белок и тот начинает оказывать влияние на внутреннее состояние клетки.
Различают два типа воздействия G-белков на клетку (рисунок ниже). В первом случае G-белки напрямую связываются с ближайшими ионными каналами и открывают либо закрывают их, что соответствующим образом меняет мембранный потенциал. Во втором случае G-белки связываются с энзимами, которые запускают работу вторичных посредников. Задействование вторичных внутриклеточных посредников ведет к многократному усилению эффективности рецепторов. Вызванные вторичными посредниками изменения являются медленными, но при этом могут глобально менять состояние всей клетки.
Работу ионотропных рецепторов принято называть быстрым взаимодействием. Для изменения мембранного потенциала требуется время порядка всего одной миллисекунды. Работу метаботропных рецепторов принято относить к медленным взаимодействиям. При вовлечении вторичных посредников изменения в клетке могут длиться от секунд до часов. Прямое управление метаботропными рецепторами работы ионных каналов протекает значительно быстрее и сопоставимо по времени с быстрым взаимодействием.
Если посмотреть на метаботропный рецептор ближе, то окажется, что у него есть семь трансмембранных доменов и два свободных конца (рисунок ниже).

Структура метаботропного рецептора
За счет свободных концов соседние рецепторы могут соединяться, создавая димеры (рисунок ниже). Димеры, в свою очередь, объединяясь, формируют рецептивные кластеры. Аминный и карбоксильный концы рецепторов выполняют роль своеобразных «застежек-липучек», которые за счет электростатического «прилипания» могут формировать различные по составу кластеры рецепторов. Так как в биологических системах нет ничего случайного, то можно предположить, что образование кластеров из различных метаботропных рецепторов несет определенный смысл. Если допустить, что кластер рецепторов реагирует на нейромедиаторы не по отдельности, каждым рецептором самостоятельно, а как единый механизм, то такая реакция может быть сопоставлена с детектированием определенных комбинаций веществ, образующихся при спиловере соседних синапсов.
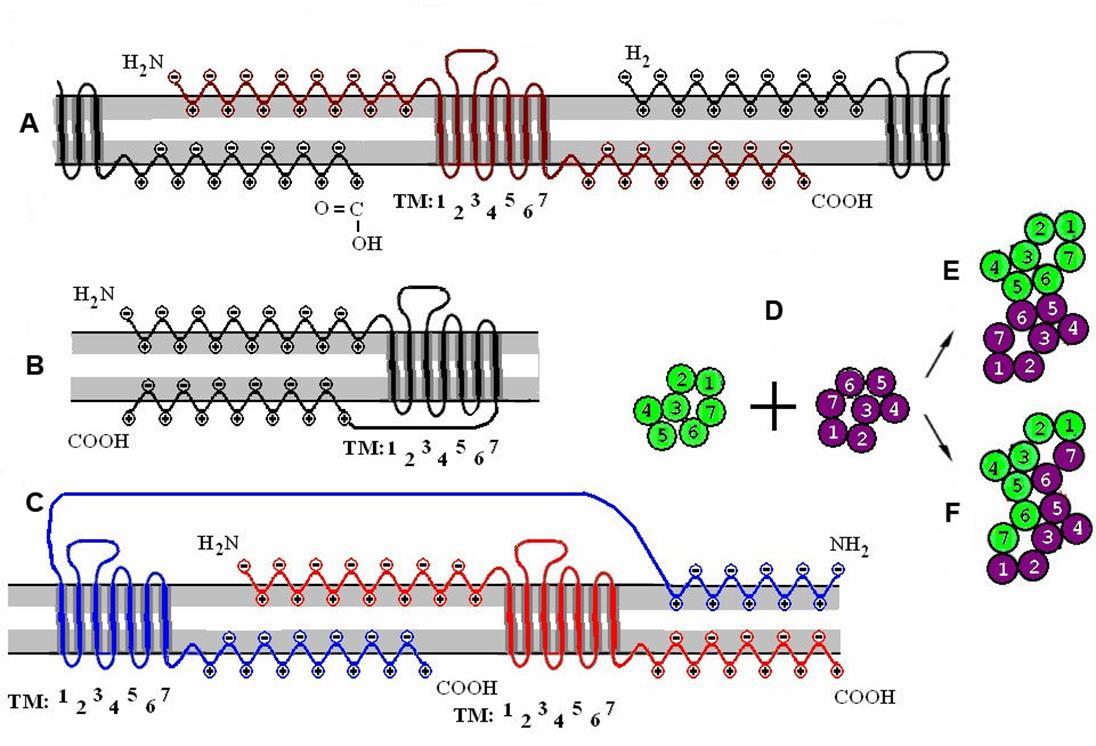
Кластеризация рецепторов. A – одиночный рецептор и его взаимодействие с окружающими рецепторами. B – мономерная рецептивная молекула. C – рецептивный димер. D – объединение двух мономеров в контактный (E) и комбинационный (F) димеры. (Радченко, 2007)
Выше описаны далеко не все механизмы, участвующие в работе нейрона. Но уже этого достаточно, чтобы осознать, что реальный нейрон не просто значительно сложнее, чем его формальный аналог. Реальный нейрон -это нечто совсем другое. Похоже, что нейронные сети — это изобретение человека, не имеющее прямых аналогов в природе. Когда искусственной нейронной сети удается решать какие-либо практические задачи, то, похоже, что аналогии с мозгом стоит проводить не на уровне нейронов и связей, а на уровне самих алгоритмических принципов, которые реализует эта сеть.
Вернемся к клеточным автоматам и вопросу о возможной биологической аналогии. Чтобы претендовать на роль элемента, участвующего в передаче информации, кандидат должен удовлетворять нескольким требованиям:
- У кандидата должны быть различимы, как минимум, два состояния;
- Должна присутствовать возможность передавать информацию о своем состоянии соседям;
- Должен существовать механизм, позволяющий кандидату под воздействием узора, созданного активностью соседей, менять свое состояние;
- Должен существовать механизм, позволяющий избирательно реагировать на различные окружающие узоры;
- Передача информации должна быть достаточно быстрой, чтобы соответствовать ритмам работы мозга;
- Так как предполагается, что паттерно-волновой механизм должен каждый раз вовлекать в передачу большое количество элементов, то энергетические затраты каждого элемента должны быть минимальны.
В разное время я рассматривал разных кандидатов на роль биологических аналогов. Обилие механизмов, свойственных мозгу, позволяет почти для всего, что есть в коре придумать гипотетическое обоснование почему именно это может быть аналогом элементов клеточного автомата. Сейчас я склоняюсь к тому, что наиболее подходящий кандидат – это тонкие веточки дендритных деревьев.
Веточки дендритных деревьев, безусловно, являются составной частью нейронов и участвуют в общем механизме его работы. Но это не мешает им проявлять и индивидуальные свойства и быть в каких-то ситуациях автономными элементами.
Когда на веточке возникает миниатюрный возбуждающий постсинаптический потенциал, он как по кабелю распространяется в пределах длины этой веточки. Можно предположить, что распространение электрического сигнала провоцирует минимальное испускание нейромедиаторов из каждого синапса, принадлежащего этой веточке. При этом испускание не затрагивает мембранный потенциал дендрита, а в основном распространяется за пределы синапса. Со стороны это будет выглядеть, как постоянная утечка нейромедиаторов. Состояние, в котором по дендриту пробегает электрический сигнал можно назвать активным состоянием элемента. В момент активности веточки дендрита вокруг нее, предположительно, создается облако нейромедиаторов. В каждом месте этого облака состав коктейля индивидуален и определяется ближайшими синапсами.
В каждом месте коры соседствуют порядка десятка синапсов от различных дендритов. Если несколько дендритов оказываются активны одновременно, то в определенных местах возникает специфический для этого сочетания нейромедиаторов коктейль. Если в таком месте окажется дендрит с метаботропным рецептором чувствительным именно к этому коктейлю, то такой дендрит может получит возбуждающий потенциал и перейти в активное состояние.
В принципе, из такого конструктора несложно собрать биологический аналог нашего клеточного автомата. За счет того, что речь идет о миниатюрных постсинаптических потенциалах и квантовом испускании нейромедиаторов энергетика такой передачи будет крайне невысока.
В клеточном автомате для создания уникальных узоров требовался случайный начальный выбор состояний и память элементов автомата относительно знакомых им узоров. Это проистекало из того, что изначально клеточный автомат был чист и однороден. Для возникновения любой повторяемой неоднородности автомату требовалась случайность и память. С дендритами ситуация несколько интереснее. Дендритные ветки изначально сильно переплетены, причем совершенно случайным образом. Такая неоднородность, по сути, уже является готовой памятью. Эта память позволяет воспринять любой сигнал и выдать предопределенный структурой переплетений ответ. Причем, ответ повторяемый. Это как хеш-функция, которая выдает результат, возможно, не очень понятный, но зато всегда один и тот же для одного и того же входного сигнала.
Упрощенно это выглядит так. Создадим в локальной области узор из нескольких веточек. Где-то в объеме этой локальной области найдутся места, в которых эти веточки пройдут рядом друг с другом. Нейромедиаторы испущенные из них создадут «коктейли». Если рядом с «коктейлями» окажутся дендритные веточки, на которых в этом месте окажется соответствующий рецептор, то такая ветка активируется.
То есть сама система случайных переплетений уже содержит внутри себя механизм создания продолжении для любых комбинаций активности. Это удобно, так как, потенциально, не требует дополнительной памяти кроме того, что уже заложено в хаос переплетений. Но такая конструкция обладает только локальной временной устойчивостью. Если конфигурация дендритов или шипиков изменится, то могут помяться и все получающиеся узоры. Можно предположить, что если мозг, действительно, выбрал такой механизм, то должны существовать системы, обеспечивающие стабильность используемых узоров, оптимизацию их распространения и минимизацию вероятности ошибок распространения. Не исключено, что изменения в дендритных деревьях и изменения, происходящие с количеством и формой шипиков – это отголоски такой оптимизации.
Чтобы проиллюстрировать описанные предположения Антон Морозов сделал 3D масштабную модель в которой воспроизвел волны, основанные на узорах из дендритных веточек. В модели веточки были заменены тонкими трубочками длиной по 50 мкм, что соответствует средней длине дендритной веточки. При той же плотности укладки дендритов, что и в реальной коре получилось нечто, изображенное на рисунке ниже.

Задается начальный компактный узор из веточек. В модели у веточек нет собственной памяти. Активируются те веточки, для которых это диктует геометрия случайных связей. Соответственно, любой случайный узор из активных веточек порождает предопределенный геометрией узор продолжения. Новый узор порождает следующий и так далее. Ниже приведено несколько шагов моделирования.





Не стоит искать в описанном механизме распространения дендритных волн какого-либо глубокого смысла, связанного с обработкой информации. По сути, мы просто показали возможный механизм передачи дискретной информации по пространству коры и между структурами мозга. Он, кстати, по своей идее аналогичен механизму передачи цифровой информации по шинам данных, используемым в компьютерах. Функция шины данных — передать узор, составленный из нулей и единиц, всем узлам компьютера. С шиной данных несколько проще, ее узор одинаково выглядит в любом месте шины. Но, теоретически, можно представить компьютер, в котором битовый сигнал на шине данных будет меняться по мере движения от одного узла к другому. Если при этом будет соблюдаться однозначное соответствие получаемых кодов, то несложно приспособить узлы компьютера для работы с такой информацией. Но не стоит и недооценивать полученную модель. Далее мы покажем, что развитие этой модели дает удивительные результаты.
Алексей Редозубов
Логика сознания. Вступление
Логика сознания. Часть 1. Волны в клеточном автомате
Логика сознания. Часть 2. Дендритные волны
Логика сознания. Часть 3. Голографическая память в клеточном автомате
Логика сознания. Часть 4. Секрет памяти мозга
Комментарии (52)


Thing-man_676
31.08.2016 14:25Кстати, довольно неплохо описано. Людям с образованием, не имеющим ничего общего с биологическим или медицинским, конечно, сложно будет что-то понять, но полезно будет узнать))

tempik
31.08.2016 14:25+2Великолепная подача информации о работе мозга! И заслуживающая внимания теория!
Пожалуйста, продолжайте публикации!

AlekseiMorozov19730316Ru
31.08.2016 14:25>… передать узор… всем узлам…
То есть реализуется широковещательный broadcast паттернов с целью поиска реципиентов «смысла». Реципиенты в свою очередь порождают новые волны поиска. Вроде бы особо «удивительных результатов» от этого не просматривается…

AlexeyR
31.08.2016 14:34Пока, удивительного ничего нет. Сам механизм вещания, хотя, и нов применительно к описанию мозга не несет чего-то революционного с точки зрения алгоритмов обработки информации. Все интересное будет дальше.
AlekseiMorozov19730316Ru
31.08.2016 15:50>… механизм вещания…… нов…
Насколько понимаю, в академических кругах «a segment of a dendrite» как «functional unit» для «dendritic spike» — вопрос уже проработанный. То есть сегменты дендритов образуют среду широковещательного распространения паттернов. Этот broadcast паттернов находит ассоциации, которые объединяются в смысловой поток.AlexeyR
31.08.2016 16:45Сегменты дендритов рассматривают, как автономные детекторы совпадения аксонных сигналов. Далее говорят от интеграции сигналов от дендритов одного нейрона (дендритных спайков) и условиях возникновения аксонного потенциала действия. Это совсем другое, отличное от механизма вещания. Классический подход — это попытка найти в нейроне следы пусть и сложного, но порогового сумматора.
AlekseiMorozov19730316Ru
31.08.2016 17:01ОK. Если это так, то это так. Но «классические» принципы работы мозга — это не только «образы», но и «ассоциации». Так что, опять таки — тут мало что нового…
AlexeyR
31.08.2016 18:43Вы совершенно правы. Пока тут нет ничего, что говорит о методах работы с информацией. Все интересное будет позже.
AlekseiMorozov19730316Ru
31.08.2016 19:23+1Полистав Ваш черновик, к сожалению, вынужден заметить, что как-то он громоздок и сыроват, некоторые вещи — явно притянуты за уши. Не говоря уже о параллелях с «квантовыми» системами — это вообще лишнее. Может быть, стоит сделать два варианта: простой, без формул, и расширенный — с формулами. А так… что можно сказать… всё суховато и не особо интересно для широкой публики…
AlexeyR
31.08.2016 20:21Для перевода выложен не черновик этого цикла, а достаточно сухая научная статья, хотя и о том же самом. Здесь на хабре совсем другой текст — живее и без формул :)
AlekseiMorozov19730316Ru
31.08.2016 20:48Старый вариант Вашей статьи, разумеется, читал пару лет назад. Будем надеяться, что в итоге Вам удастся отказаться от сложных ответов на простые вопросы. Всё устроено просто.
kennivich
02.09.2016 02:08Слежу за развитием вашей модели с большим интересом, но одна проблема постоянно бросается в глаза и я не нашел ее решения ни в этих статьях, ни в той, что еще переводится. Чтобы волны могли приходить и уходить в любые стороны, аксоны и дендритные деревья нейронов должны покрывать хотя бы частично объем по всем направлениям от клетки (2). Для дендритов это представить довольно легко, но аксоны обычно образуют синапсы на некотором расстоянии от дендритного дерева (1).

Красным активный нейрон и его дендриты, зеленым его аксон, серым соседние нейроны.
Даже в отображении на плоскости сложно представить, как аксон мог бы иметь контакты с нейронами по все стороны от сомы, а в трехмерном пространстве это выглядит еще страннее. Условно говоря, сложно представить как нейрон мог бы передать сигнал и направо и налево, при том что основная ветвь аксона уходит в одну сторону.AlexeyR
02.09.2016 02:15Аксон очень сильно ветвится, образуя коллатерали. Ветвление достаточно равномерно покрывает ближайший к нейрону объем, затем аксон может уйти очень далеко. Есть работа, где были изучены под оптическим микроскопом порядка 15 пар приблизительно соседних нейронов коры. В среднем у них оказалось по 7 общих синапсов.
AlexeyR
02.09.2016 02:20Кстати, рекоммендую: «Valentino Braitenberg. Almut Schuz
Cortex: Statistics and Geometry of Neuronal Connectivity» — эта книга есть в сети. Там такие вопросы разобраны очень подробно.
shabanovd
02.09.2016 10:24Также передача волны идентификатора происходит только дендритами, но такой же механизм кода идентификатор распространяется по аксонам (сам идентификатор формируется несколькими аксонами).
rodgar
31.08.2016 14:34Хм. Возвращаясь к игре «Жизнь», получается есть два варианта
1. плотное размещение клеток, но правила активации различаются от клетки к клетке, и эволюционируют со временем.
2. правила едины для всех но кубик разряженный, с пустотами, и эволюционирует расположение клеток.
А при смешении этих вариантов на одном поле и начинается магия. Если я всё правильно понял.AlexeyR
31.08.2016 14:36Неравномерность, созданная клубком связей, облегчает создание уникальных узоров. Но магии не несет. Магия будет позже и совсем в другом.

Lucian
31.08.2016 14:59Алексей, спасибо за труды, скажите пожалуйста, почему на картинке (желтая в художественном стиле), аксон разветвляется, т.е. по сути крепится к самому нейрону и к его дендритам, влияет ли это каким-то образом на восприимчивость нейрона к входящим сигналам? Просто эта картинка наводит на некоторые сомнения в традиционных сетях, где нейрон нарисован схематически и имеет только 1 вход от 1-го нейрона.
AlexeyR
31.08.2016 15:11Если взять два реальных соседних нейрона, то в среднем у них будет около 7 общих аксоно-дендритных синапсов. Синапсы на теле нейрона составляют, где-то, пять процентов от общего числа. Есть шанс, что общий синапс окажется и на теле нейрона. Причем, число и место совместных связей выбирается нейронами случайным образом, как получится. Это слабо похоже на связи с весами из идеологии нейронных сетей.
Вывод, который, можно сделать — это, что нейронные сети и мозг реализуют сосвсем разные принципы и мозг совершенно не похож на нейронные сети. Далее я расскажу, гипотезу о том, что действительно делает мозг.AlekseiMorozov19730316Ru
31.08.2016 16:26>… мозг совершенно не похож на нейронные сети…
Не совсем удачное выражение. Биологические нейронные сети и искусственные «нейронные» сети — разные понятия. К тому же, это лишь вопрос типа и качества используемой модели.AlexeyR
31.08.2016 16:51Да, это разные понятия и я, по возможности, стараюсь уточнять о каких сетях идет разговор. Относительно типа и качества модели, то это подразумевает набор принципов по которым функционирует сеть. Вот тут есть сложность. Существующие нейронные сети, с моей точки зрения, которую я попробую обосновать, построены на принципах кардинально отличных от принципов биологических нейронных сетей.
AlekseiMorozov19730316Ru
31.08.2016 17:12Уровень Вашего нахождения «в теме», видимо, уже близок к уровню нейробиологов. Вопрос лишь в том, что, собственно, это даёт… В любом случае, «одержимость» Вашего мозга мозгом как таковым достойна похвалы…
wvitus
31.08.2016 18:43Должен существовать механизм, позволяющий избирательно реагировать на различные окружающие узоры;
Если для наблюдения за глобальными узорами, разбить все пространство на области, доступные наблюдению единичным нейроном и организовать из них сеть, то это ж уже будет базис для самосознания, ага?
RockPresident
02.09.2016 18:03Сейчас я склоняюсь к тому, что наиболее подходящий кандидат – это тонкие веточки дендритных деревьев.
Можете квантифицировать слово "склоняюсь"? По вашей интуиции (предполагая что клеточный автомат в мозге действительно есть) — каков шанс что он реализуется именно дендритными веточками, и какой шанс что чем-то ещё?
AlexeyR
02.09.2016 18:49То, что общий принцип клеточного автомата (с волнами паттернов) свойственен мозгу я практически уверен. Далее я приведу очень сильные доводы в пользу этого. Что аналог элементов — дендритные веточки, вероятность того, что я прав — процентов 60. Опять же, далее я покажу многое, что говорит в пользу веточек.
AlekseiMorozov19730316Ru
02.09.2016 19:23>… общий принцип клеточного автомата (с волнами паттернов) свойственен мозгу…
В Вашем старом варианте от 2014 года (http://arxiv.org/abs/1406.6901) были, по сути, только волны по нейронам в качестве элементов. Оставалось лишь прийти к выводу, что волна приносит к нейрону паттерн, а нейрон, как ему и положено, на этот паттерн реагирует. Если паттерн опознан как свой, то после активации всего нейрона, дендриты, присоединённые к его аксону, порождают ещё и волну с паттерном этого нейрона. В этом и состоит простая и понятная суть широковещательного распространения паттернов с целью их ассоциирования. Не надо ни «автоматов», ни «голограмм», ни ещё не пойми чего…
AlexeyR
02.09.2016 20:59Части первая по четвертую — это, по сути, вступление. В них нет чего-то принципиально нового в сравении с 2014 годом, скорее, так, уточнения. Все самое интересное будет дальше. Вот там и станет понятно, зачем все эти сложности и голограммы.
Клеточный автомат в описании появился из-за того, что я его запрограммировал и отвечаю за его работоспособность. А, вот, относительно мозга я могу только строить гипотезы. Мне показалось, что лучше четче разделить факты и предположения.

VDG
03.09.2016 22:36Дендритные спайки могут возникать и непосредственно в дендрите. Для этого требуется, чтобы в течении короткого временного интервала (порядка 3-10 мс) на небольшом участке дендрита случилось достаточно много синаптических возбуждений.… Добиться синхронной активности четверти всех синапсов можно при искусственном возбуждении «в пробирке». Трудно сказать возможно ли такое в живой ткани.
Из-за чего тогда у нейронов появляется ПД, если дендритные спайки столь маловероятны?AlexeyR
04.09.2016 01:12В кабельной теории дендритов показывается, что токи, возникающие в дендритах могут складываться и доходить до сомы. Глиальные клетки могут модулировать количество выделяемого синапсом нейромедиатора. Метаботропные рецепторы могут открывать ионные каналы и создавать возбуждающие потенциалы.
Всего, что вляет на работу нейрона много. Трудно сказать, что именно вносит больший вклад. Мои соображения по этому поводу будут в 4 части.AlekseiMorozov19730316Ru
04.09.2016 08:07>… токи, возникающие в дендритах могут складываться и доходить до сомы…
Видимо, самым простым кандидатом для среды широковещательного распространения выходных паттернов нейронов всё-таки являются именно «заряженные частицы», распределённые по нейронам, а не сами сегменты нейронов. Тогда всё встаёт на свои места. Спайк возбуждённного нейрона не только передаёт возбуждение связанным с ним нейронам, но и модулирует выходным паттерном нейрона несущие электрические колебания в отделах мозга. При этом, в общем случае, выходные паттерны активных нейронов могут накладываться друг на друга, и распространяется уже некоторый суммарный паттерн. Если распространившийся до нейрона паттерн «совпал» с входным паттерном этого нейрона, то демодуляция несущих электрических колебаний в дендритах нейрона вызывает активацию всего нейрона, то есть, в соответствующих терминах, потенциал действия. Таким образом в самом простом случае обеспечивается «ассоциирование» выходного паттерна одного нейрона с входным паттерном другого нейрона. Но возможно и «ассоциирование» выходных паттернов нескольких нейронов с одинаковым входным паттерном нескольких других нейронов. Нейрон, обладая естественными входным и выходным паттернами, связан «ассоциациями» с другими нейронами. Входной паттерн нейрона определяется геометрией его дендритного дерева, а выходной паттерн — геометрией дендритов, присоединённых к его аксону.AlexeyR
04.09.2016 14:09Есть несколько доводов, которые очень сильно говорят в пользу дендритных веточек. Они будут описаны в четвертой части.
AlekseiMorozov19730316Ru
04.09.2016 17:26По сути, совокупность сегментов дендритов нейрона определяет своей геометрией входной паттерн этого нейрона, а совокупность сегментов дендритов, присоединённых к аксону нейрона определяет своей геометрией выходной паттерн этого нейрона. Нет никакого смысла наделять сегмент дендрита функциональностью целого нейрона. Понятно, конечно, что широковещательное распространение паттернов «ломает» идею Вашего «автомата», но это «сreative destruction» — такова суть процесса поиска истины…
AlexeyR
04.09.2016 20:01Вы несколько в своих фантазиях и говорите о модели с совсем другими свойствами. Позже станет понятно зачем нужны именно те свойства, что описываются в этом автомате. Наберитесь терпения.
AlekseiMorozov19730316Ru
04.09.2016 20:27:) Да, Вы правы. Совсем разные свойства, по сути — мало что общего. Ваша модель, разумеется, более-менее понятна, понятно и резюме Вашей модели — «мозг совершенно не похож на нейронные сети». Написал в песочницу текст «Широковещательное распространение паттернов нейронов мозга». Может кого заинтересует эта идея.
unbalanced
Интригующе.
С трудом продравшись сквозь описание железной части, все же вангую направленный граф. Угадал? :)
Halt
А причем тут направленный граф?
unbalanced
Ну как же. Суть цикла статей, насколько я понял — поиск замены слоеным нейросетям.
Автор в конце говорит о пачке переплетенных возбуждающихся от контакта или proximity цилиндров. Понятно, что цилиндры форматом хранения и обработки не сделаешь, но судя по всему эта конструкция редуцируется до направленного графа. Направленный — потому что возбужденный цилиндр уже возбужден.
Вот я и спрашиваю, угадал?
Halt
Угадал или не угадал пусть автор напишет. Но у меня по всем предыдущим описаниям складывается скорее обратное впечатление.
Описанные ранее абстрактные клеточные автоматы действительно довольно хорошо ложатся на нейроны, как впрочем и на любую другую систему с двумя выраженными состояниями и дешевым, но не бесплатным переключением между ними. Это автоматически приводит к появлению волн просто потому, что у элементов есть гистерезис активации (что еще и решает проблему независимости от направления распространения волны).
В абстрактной модели важен лишь факт активации соседей. В физической же компактность упаковки элементов приведет к перевозбуждению, если все элементы будут реагировать только на один стимул. Логично предположить, что в условиях биологического разнообразия среды это решается тем, что стимулы оказываются разные.
Ну а дальше все по законам автоматов: две волны, пришедшие с небольшим интервалом времени оказываются проассоциированы в этой области. Если одной волны достаточно для превышения порога активации, может произойти «выборка» по «ключу», и т. д.
Самое интересное на мой взгляд во всей этой модели то, что ее эволюция может быть легко описана. Легко представить и выделить некоторые отдельные этапы развития системы которые приводили к качественному росту и закреплялись, что в конечном итоге привело к подобной организации сети.
На мой взгляд это выглядит куда более правдоподобно, чем альтернативные теории, которые приходилось слышать.
P.S.: Видео, представленное в статье дает понять, что нейроны растут в общем-то произвольно и синапсы образуются там где им удобно. В противовес другой теории, когда нейроны «вынюхивают» и целенаправленно тянутся к желаемой точке интеграции. Хотя и это поведение можно описать, если считать что нейрон стремится оптимизировать себя и свести энергозатраты к минимуму при сохранении связности.
DrZlodberg
Выглядит действительно весьма правдоподобно.
Однако если берём за основу клеточные автоматы, то (если не использовать автоматы в многомерном пространстве) скорее всего всё-таки граф. Собственно обычные 2D автоматы вполне заменяются регулярным графом. Что проще, граф или многомерный автомат — вопрос.
Кстати и нейросети вполне можно представить графом с весами. Хотя это уже предыдущему комментарию ответ :)
unbalanced
Т.е. можно входной слой с, например, тысячей нейронов, принимающих каждый свой пиксель, схлопнуть в один?
DrZlodberg
Про «схлопнуть» не понял. Однако в контексте треда каждый нейрон вполне можно заменить узлом графа.
unbalanced
Это я ваш коммент недопонял)
Так-то да, согласен.
AlexeyR
Нет, будет не граф. Причем, далеко не граф. Угадать, что будет дальше из того, что рассказано очень непросто. У меня на это ушел не один год.
unbalanced
Интересно будет узнать)
Shortki
Развитой плотно связанный граф формирует модель пространства с довольно интересной метрикой зависящей от плотности связности. Я бы копал здесь.