
Мы остановились на том, что лактозный оперон похож на логический элемент AND. Но откуда у него берутся цифровые свойства? Ведь оба входных сигнала (концентрации цАМФ и лактозы), вообще-то, аналоговые. Попробуем нарисовать входные функции лактозного оперона.
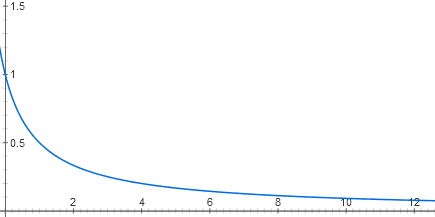
Каждый вход лактозного репрессора состоит из двух этапов молекулярного узнавания. Лактоза связывается с лактозным репрессором, а репрессор — с ДНК. В клетке находится несколько десятков молекул репрессора, каждая из которых, пока нет лактозы, может связываться с началом лактозного оперона. Когда она там связана, РНК-полимераза не может начать работу. Из-за теплового движения молекулы репрессора то и дело отваливаются от ДНК и присоединяются обратно. Если репрессора нет совсем, оперон работает на полную мощность (сейчас для простоты считаем, что катаболический активатор есть в избытке). При малых концентрациях репрессора он снижает активность оперона практически линейно. Но дальше эффект каждой новой порции репрессора все меньше и меньше, и в целом график зависимости близок к гиперболе y = 1 / (x + 1)
Например, у нас есть реакция, в которой из веществ А и В образуется вещество С. Реакция обратима, то есть С может распадаться обратно на А и В в тех же самых условиях:
A + B <-> C
В колбе, где идет эта реакция, установится какое-то равновесие между прямым и обратным направлением реакции. В системе будут все три вещества А, В и С в каких-то концентрациях (в физической химии концентрацию вещества принято обозначать его символом в квадратных скобках, [A], [B] и [C] в нашем примере). Закон действующих масс связывает равновесные концентрации веществ между собой: произведение концентраций продуктов, деленое на произведение концентраций исходных веществ, постоянно.
[C] / ([A] * [B]) = K
Буквой К здесь обозначена константа равновесия реакции. Она зависит только от химической природы реакции (т. е. какие вещества реагируют) и от температуры. Концентрации веществ, давление, катализаторы и прочие факторы не влияют на константу равновесия.
Например, если мы добавим в колбу побольше исходного вещества А, то равновесие сдвинется в сторону вещества С, его концентрация возрастет, а концентрация вещества В (которого мы не добавляли) при этом упадет. Однако отношение [C] к ([A] * [B]) останется постоянным.
Итак, у нас есть обратимая реакция связывания активной формы репрессора с опероном: RepA + OpA <-> Op (связывание репрессора превращает активную форму оперона в неактивную). Она имеет свою константу равновесия К{R-O} = [Op] / ([OpA] * [RepA]). Нам надо получить отсюда зависимость [ОрА] (количества активной формы оперона) от [RepA] (концентрации репрессора). Еще мы знаем, что в клетке только один лактозный оперон, который может быть в активной или неактивной форме: [Op] + [OpA] = 1
Из второго уравнения выражаем [Op] через [OpA] и подставляем в первое уравнение:
К{R-O} = (1 — [OpA]) / ([OpA] * [RepA])
После преобразований получаем:
[OpA] = 1 / (K{R-O} * [RepA] + 1)
То есть функция имеет вид y = 1 / (x + 1)
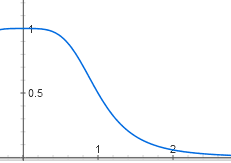
Молекулы репрессора плавают в клетке и связываются с ДНК только пачками по четыре. Чтобы активная форма репрессора превратилась в неактивную, она должна связаться с четырьмя молекулами лактозы — одной или двух недостаточно, надо по одной молекуле лактозы на каждую белковую субъединицу счетверенного репресора. Пока лактозы в клетке мало, ее молекулы связываются с репрессором по одной-две и он остается активным. Но после некоторой пороговой концентрации лактозы большинство молекул репрессора связывают по четыре молекулы лактозы и переходят в неактивную форму. График активности репрессора в зависимости от количества лактозы поэтому имеет S-образную форму и описывается функцией вида y = 1 / (1 + x^4).
RepA + 4 Lac <-> Rep
В уравнении для константы равновесия в этом случае появляются степени:
К{R-L} = [Rep] / ([RepA] * [Lac]^4)
Опять мы используем тот факт, что суммарная концентрация обоих форм репрессора постоянна:
[Rep] + [RepA] = n, где n ? 50
И получаем, что [RepA] = n / (К{R-L} * [Lac]^4 + 1)
Теперь, чтобы получить входную характеристику оперона по лактозе, надо подставить вторую функцию в первую. Она будет с точностью до коэффициентов иметь такой вид:
y = 1 / (1 + (1 / (1 + x^4)))
Ее график тоже S-образно изгибается. Получается, что лактозный оперон не реагирует на малые концентрации лактозы. При достижении пороговой концентрации появляются неактивные молекулы репрессора, связавшие по четыре лактозы, и оперон довольно резко включается.
Второй регуляторный вход лактозного оперона тоже состоит из двух шагов молекулярного узнавания: цАМФ связывается с активатором, а активатор — с ДНК. Отличие от лактозного входа в том, что здесь связывание молекул (активатора или цАМФ) повышает активность того, с чем они связались (оперона или активатора). То есть графики будут выходить из нуля и приближаться к горизонтальной прямой где-то выше оси Х.
2 цАМФ + Akt <-> AktA
Константа равновесия K{Ц-А} = [AktA] / ([Akt] * [цАМФ]^2)
Опять мы пользуемся тем, что сумма концентраций [AktA] и [Akt] постоянна, так как в клетке есть около 100 молекул активатора, переходящих между активной и неактивной формой. Выражая [Akt] через [AktA], получаем:
K{Ц-А} = [AktA] / ((100 — [AktA]) * [цАМФ]^2)
[AktA] = K{Ц-А} * [цАМФ]^2 / (1 + K{Ц-А} * [цАМФ]^2)
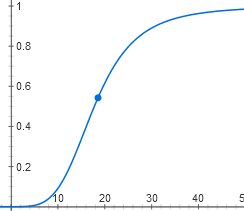
То есть зависимость [AktA] от [цАМФ] имеет вид y = x^2 / (1 + x^2)
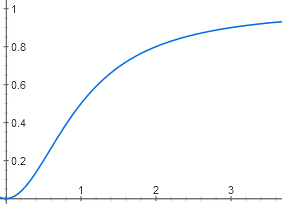
Из-за связывания двух молекул цАМФ график выглядит не как гипербола, а как S-образная кривая. Хотя пороговый эффект на ней заметен слабее, чем на графике для лактозы, потому что степень не четвертая, а только вторая.
Зависимость активности оперона от количества активатора имеет другой нюанс. Если связывание репрессора подавляет оперон почти полностью (где-то до 0,1%, что ниже погрешности биохимических измерений), то отсутствие активатора подавляет активность только до 5%. Поэтому график будет выходить не из нуля, а из точки (0; 0,05):
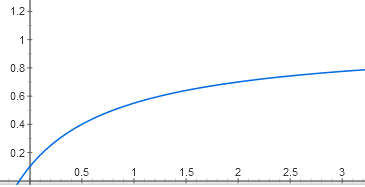
y = 0,05 + (0,95 * x / (1 + x))
Активность = [OpA] + 0,05 * [Op]
Зависимость [OpA] от [AktA] мы можем легко вывести по аналогии с предыдущими тремя. Активатор связывается по одной штуке, значит, [AktA] будет входить в формулу в первой степени. Он активирует оперон, значит, [AktA] будет и в числителе, и знаменателе:
[OpA] = K{А-O} * [AktA] / (K{А-O} * [AktA] + 1)
То есть функция имеет вид y = x / (1 + x)
И это только вклад оперона, связанного с активатором! Чтобы учесть вклад оперона без активатора, надо добавить слагаемое (1 — эта дробь) * 0,05. К счастью, оба слагаемых имеют общий знаменатель, поэтому функция усложняется совсем немного:
y = 0,05 + (0,95 * x / (1 + x))
Осталось получить зависимость активности оперона от концентрации цАМФ. Для этого надо подставить первую формулу (для концентрации активного CAP в зависимости от концентрации цАМФ) во вторую формулу (для активности оперона от концентрации активного САР)…
… И получить четырехэтажную дробь:
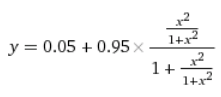
У нас получилась самая сложная функция из всех в этой статье. Впрочем, ее график выглядит похоже на график более простой зависимости активного CAP от концентрации цАМФ, y = x^2 / (1 + x^2):
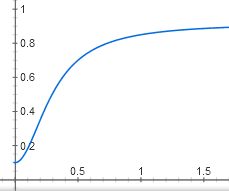
Тоже S-образная кривая с некоторым пороговым эффектом, которая медленно приближается к горизонтали y = 1. Только начинается не из нуля, а из точки (0; 0,05).
Осталось понять, как взаимодействуют два входа. В случае лактозного оперона ответ простой — никак. Лактозный репрессор и катаболический активатор не влияют на связывание друг друга с ДНК. Поэтому связывание этих двух белков можно считать независимыми событиями. Полная активность оперона достигается, когда и активатор связан, и репрессор не связан. Вероятность такого совпадения равна произведению вероятностей каждого из них по отдельности. Значит, чтобы получить функцию активности лактозного оперона от двух переменных ([Lac]) и ([цАМФ]), надо просто перемножить функции от каждой этой переменной:
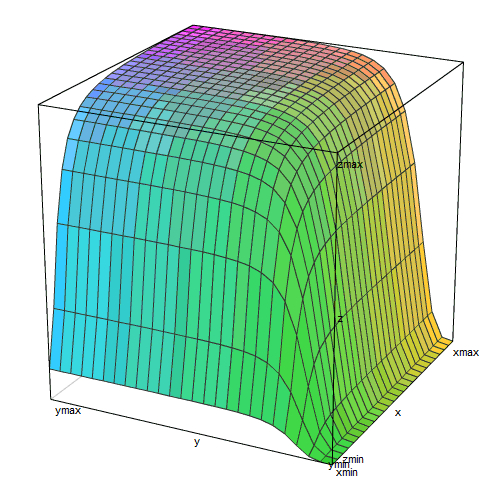
Активность = (1 / (1 + (1 / (1 + [Lac]^4)))) * (0,05 + 0,095 * ([цАМФ]^2 / (1 + [цАМФ]^2)) / (1 + ([цАМФ]^2 / (1 + [цАМФ]^2))))
Трехмерный график этой функции
похож на плато, обрывающееся ущельями по двум сторонам. Правое ущелье (малые концентрации лактозы) более глубокое и плоскодонное, чем левое (малые концентрации цАМФ).
В реальной жизни кишечной палочки вокруг нее лактозы либо нет совсем (чаще всего), либо ее концентрация выше пороговой и лактозный репрессор практически не мешает работе оперона (когда хозяин съел что-то молочное). Концентрация цАМФ — это внутренний сигнал, который вырабатывается самой клеткой. Она тоже бывает либо слишком малой для включения лактозного оперона (когда есть глюкоза или крахмал), либо достаточной для включения на 95% и более (если ничего вкуснее лактозы нет). То есть почти всегда лактозный оперон находится в условиях либо плато на этом графике, либо в одном из ущелий.
Входные функции лактозного оперона измерены экспериментально. Способ измерения напоминает прием отладки микроконтроллеров «повесить светодиод на выходную ногу». Берется регуляторная область оперона и соединяется с геном зеленого флуоресцентного белка. Эта генная конструкция вставляется в клетки кишечной палочки, из которой удаляется штатный лактозный оперон (чтобы концентрация лактозы была жестко задана экспериментатором) и штатная система, кодирующая голод уровнями цАМФ. После этого активность лактозного оперона можно точно измерять спектрофотометром по уровню зеленой флюоресценции. Оказывается, что в пределах точности измерений (1%) теория полностью сходится с экспериментом).
Дно левого ущелья на графике входной функции (при малых концентрациях цАМФ) находится выше нуля. Это не баг, а фича: благодаря ей при наличии нескольких сахаров сразу, кишечная палочка оказывается готова быстро переключиться на лактозу, когда глюкоза кончится. Мутанты кишечной палочки, у которых регуляторная функция лактозного оперона ближе к чистому AND (то есть без цАМФ оперон не работает), переключаются с глюкозы на лактозу по часу и более: в момент исчерпания глюкозы у них нет ферментов усвоения лактозы, а значит, нет и энергии, чтобы их быстро сделать. Нормальная кишечная палочка («дикий тип») встречает этот момент, уже имея сколько-то ферментов усвоения лактозы и ей хватает 15-20 минут для полного переключения на новый сахар.
Теперь отвлечемся от лактозного оперона и посмотрим шире, на что способны логические элементы на такой химической базе. Практически везде в биохимических сигнальных системах мы встречаем молекулярное узнавание, которое описывается функциями вида y = x^n / (1 + x^n) для активаторов и y = 1 / (1 + x^n) для репрессоров. Степень n в этих функциях показывает количество связываемых однотипных молекул для получения эффекта, и бывает обычно равна 1, 2 или 4 (чаще всего 2). Можно получить более сложные логические ключи, добавляя в начало оперона больше сайтов связывания регуляторных белков. Этих белков может быть больше двух типов. Эффекты разных белков на посадку РНК-полимеразы могут складываться (OR) или перемножаться (AND). Но в целом мы будем иметь дело с какой-то комбинацией (сложением или перемножением) базовых функций y = x^n / (1 + x^n) и y = 1 / (1 + x^n) с разными степенями и коэффициентами.
Некоторые интересные функции получаются очень просто. Например, один и тот же регуляторный белок может связываться (в спаренной форме) с двумя участками промотора. В одном участке он действует как репрессор, а в другом — как активатор. Для работы оперона надо, чтобы сайт репрессора был пуст, а сайт активатора — занят. Получается функция с четким максимумом:
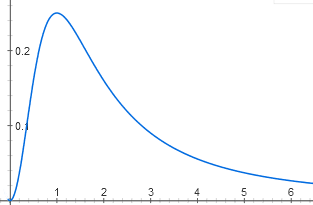
y = (1/ (1 + x^2)) * (x^2 / (1 + x^2))
Без этого белка оперон неактивен, потому что пуст сайт активатора, а при высоких концентрациях он неактивен, потому что занят сайт репрессора. Максимальная активность будет, когда и сайт активатора, и сайт репрессора заняты с вероятностью 50%.
Еще можно сделать два сайта связывания активатора, в одном активатор будет сильно связываться с ДНК, но слабо активировать ген, в другом — слабо прикрепляться к ДНК, но сильно активировать ген. В этом случае получается функция вроде y = x^4 / (1 + x^4) + 0.3 * (10x)^4 / (1 + (10x)^4), со ступенчатым графиком:
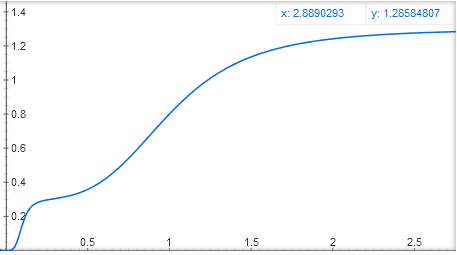
Математики, ау! Можете рассказать в комментариях, что еще можно и что нельзя получить при сложении и перемножении этих базовых функций y = x^n / (1 + x^n) и y = 1 / (1 + x^n)?
В следующей части займемся схемами из таких логических элементов.
Комментарии (11)

alex_vesna
15.12.2016 17:36Вопрос утопичный, но всё же…
Вот возьмем и заменим все биохимические элементы из статьи, на схожие из области микроконтроллеров, и подобных тем… Ну скажем пусть будет не знаю, PID для пространственного положения, он немного сложнее подобной системы, но вполне сопоставим. Предположим вы пишете код, для того что бы всё работало примерно так же как всё описано в статье… Оцените трудозатраты с нуля, на подобный законченный код, и «умственные усилия» потребные на его написание.
А теперь собственно вопрос: КАК система без разума, чисто хаотическим тыком, создает системы подобной(Я молчу про организмы в целом) сложности, как бы само собой???
Оговорюсь что любые идеи на тему инопланетян и богов, — ИМХО бред. даже не желаю рассматривать, и каменты в этих ключах, — неуместны.

QDeathNick
15.12.2016 18:24Ну не хаотический тык же, а упорядоченный и плавный. PID же тоже не сразу появился, эволюционировал.

Sormovich
18.12.2016 19:15КАК?
1) Попытки написать симулятор клетки уже есть:
http://www.nanonewsnet.ru/news/2016/uchenye-rabotayut-nad-sozdaniem-pervoi-trekhmernoi-modeli-zhivoi-kletki
2) Либо «Разум» всё же есть, как считает Андрей Николаевский. Пусть даже как гигантская программа:
http://newfiz.narod.ru/nikol/index1.html
HellMaus
19.12.2016 01:15+1С этим симулятором клетки есть две проблемы. Во-первых, если всерьез пытаться моделировать сразу много уровней биологической организации (от молекул до клетки, как у них), вычислительной мощности современных суперкомпьютеров не хватит где-то на двадцать-тридцать порядков. Во-вторых, для такой модели (и я об этом планирую писать) нужны миллиарды параметров взаимодействия молекул — каждого вещества с каждым, и не только попарные. Чтобы экспериментально измерить их все, не хватит армии в миллион китайских лаборантов
Sormovich
19.12.2016 22:32-1Согласен. Даже кластерные технологии не вытягивают. Тут либо уповать на то, что «дожмут» квантовый компьютер. Либо создадут хороший алгоритм «глубокого машинного обучения». Судя по всему, даже аналог SETI не вытянет такую задачу. Но вот что интересно: каждый уровень в биологии самоподдерживается посредством ОС (обратная связь). То есть саморегулируется. И очень жёстко. Интересно об этом у Загускина. С.Л. То есть, можно как то воздействовать на уровни организма, управляя им (в сторону выздоровления). Неплохо бы в сторону принудительной регенерации.
«Ритмы клетки и здоровье человека»
https://www.youtube.com/watch?v=wiM_zYiQ_N4
http://www.biophys.ru/lib/books/sci-books/419-zaguskin-2010
lavmax
15.12.2016 23:58Вам никогда не хотелось сделать свою клетку. Какой-нибудь простой автомат: мембрана + 2-3 оперона + такие вот регуляторы + минимальные сопутствующие механизмы. И заставить ее делать что-то, реагировать на что-то или производить что-то? Насколько это вообще реально при текущем уровне науки?
HellMaus
18.12.2016 19:18+1Вы имеете в виду реальную «мокрую» клетку или компьютерную модель?
В случае реальной клетки для работы любого оперона нужно:
1) система синтеза белков — рибосома с ее обвязкой (под 100 белков)
2) система транскрипции — РНК-полимераза и ее вспомогательные белки (еще десяток)
3) системы энергообеспечения, мембранного транспорта и еще кое-что по мелочи
На круг выходит, что минимальная клетка, которая может жить в пробирке — это 300-400 белков. У Крейга Вентера при выкидывании генов из микоплазмы примерно столько и получилось.
lavmax
Великолепная статья, спасибо!
А кто такой САР? По смыслу похоже на лактозный репрессор, но аббревиатуру расшифровать сходу не получилось :)
HellMaus
CAP — Catabolic activator protein, катаболический активатор. Через него обрабатывается сигнал голодания (цАМФ). Поправлю в тексте, спасибо!
lavmax
ОК, теперь понял.