

Люди, как и многие другие животные на планете, считаются по большей степени дневными существами. Практически все системы нашего организма настроены на активность в течение дня и на отдых в течение ночи. Самым же главным регулятором того, когда нам спать, а когда бодрствовать, является наш мозг. Он же отвечает и за обработку получаемой посредством органов чувств информации. Точность этой информации зависит от того, в каких обстоятельствах она была получена и от того, как мы ее интерпретируем: в полумраке стоит вешалка, на которой висит пальто, а наш мозг говорит нам: «это монстр, который сейчас нас сожрет». Однако при определенных обстоятельствах одни сигналы могут компенсировать недостатки других.
Сегодня мы с вами познакомимся с исследованием, в котором ученые из Мэрилендского университета (США) поместили лабораторных мышей в кромешную темноту на целую неделю. Как менялось поведение мышей во время опыта, какие изменения нейронных сетей в мозге у них происходили и какие выводы сделали ученые из своих наблюдений? Свет на эти вопросы прольет доклад исследователей. Поехали.
Основа исследования
Все мы знаем, что при переходе из хорошо освещенной в темную комнату, нашему зрению необходимо немного приспособиться. Этот процесс называется достаточно прозаично — адаптация глаз. Когда мы заходим в плохо освещенное помещение, наши глаза начинают панически приспосабливаться к новым условиям, после чего этот процесс замедляется. Если же перед входом в темноту закрыть глаза на пару минут, то адаптация будет протекать лучше и медленнее. Бытует мнение, что именно по этой причине у пиратов была повязка на одном глазу: когда они заходили в трюм корабля, где освещение было не самое хорошее, они переодевали повязку на другой глаз, тем самым ранее закрытый лучше воспринимал информацию. Это похоже на правду, но может быть и простой сказкой.
Тем не менее, кроме зрения имеются и другие чувства, в том числе и слух. Мы часто слышали, что слепые люди, якобы, лучше слышат. Также считается, что сенсорная пластичность внутри одной модальности* более развита у детей, однако даже у взрослых может развиваться кросс-модальная пластичность.
Модальность* — принадлежность определенного сигнала к определенной сенсорной системе. Существуют такие типы модальности: визуальная, аудиальная, болевая, кинестетическая и т.д.В случае кросс-модальной пластичности нейроны мозга реорганизовываются таким образом, чтобы объединить функции двух или более систем. Такое явление часто возникает на фоне сенсорной депривации (ухудшения) той или иной системы ввиду травмы или заболевания. Самая ярко выраженная нейронная реорганизация присутствует у людей, которые были слепы или глухи с самого рождения. То есть чем продолжительнее депривация, тем сильнее кросс-модальная пластичность.
Другими словами, утрата сенсорной модальности может быть компенсирована пластичностью в остальных чувствах. Ранее проведенные исследования показали, что слепые с рождения люди значительно лучше воспринимают звуковые сигналы, местоположение их источников (локализация звуков) и частотные характеристики. Очевидно, что подобные изменения более развиты в случаях врожденных сенсорных дефектов. Однако даже у взрослых особей различных видов может развиться сенсорная и кросс-модальная пластичность ввиду изменений окружающей среды.
К примеру, короткие периоды визуальной депривации у грызунов на клеточном уровне увеличивают частотную селективность в слуховой коре мозга (А1), снижая при этом пороговое значение и увеличивая скорость передачи импульсов нейронами L4 (thalamorecipient layer 4) слуховой коре мозга. Подобные изменения на клеточном уровне являются результатом изменений нейронных цепей. Воздействие темноты приводит к усилению таламических входящих сигналов в L4, восходящих внутрикортикальных связей от нейронов L4 до нейронов L2/3 (thalamorecipient layer 2/3), а также к усилению возбуждающих и ингибирующих внутрислойных связей в пределах L2/3, межслойных восходящих соединений от L4 до L2/3 и соединений обратной связи от L2/3 до L4.
Однако сенсорные стимулы преобразовываются в информацию не отдельными нейронами, а их группами (популяциями). Потому изменение связи между нейронами внутри таких популяций может иметь непосредственное влияние на формирование информации из входящих сигналов.
Поскольку восприятие звуковых частот различается между L4 и L2/3, ученые решили проверить, может ли временное воздействие темноты на особей зрелого возраста реструктурировать нейроны и их связь друг с другом внутри слуховой коры мозга лабораторной мыши. Для наблюдений за процессом изменений в мозге была использована двухфотонная кальциевая (Ca2+) визуализация. Были проведены измерения вызванной звуком активность нейронов L2/3 и L4, показавшая наличие повышенной частотной избирательности.

Изображение №1: двухфотонная кальциевая (Ca2+) визуализация нейронов в A1 у мышей.
В роли подопытных выступили лабораторные мыши, которых случайным образом разделили на две группы: контрольную (жили при нормальном освещении, 9 особей) и тестовую (жили в темноте, 6 особей). Пребывание в темноте (DE — dark exposure) было относительно недолгим — 1 неделю (1A).
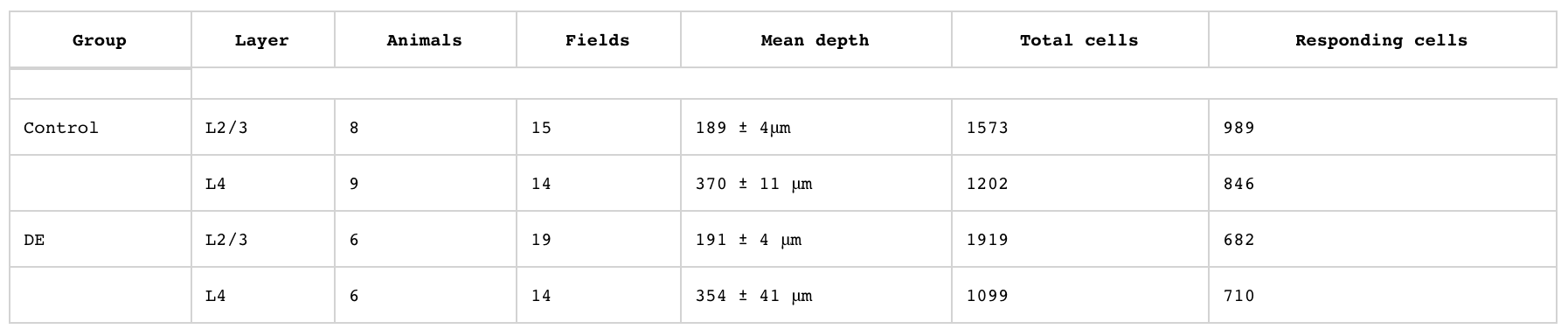
Таблица №1: кальциевая визуализация позволила получить картину активности сотен нейронов в каждом из слоев слуховой коры мозга.
Ученые также отмечают, что ранее проведенные исследования показали, что DE не вызывает каких-либо изменений в свойствах внутренней проницаемости клеток (1C). Это крайне важно, поскольку во время визуализации использовался кальциевый индикатор GCaMP6 (1В).
Чтобы охарактеризовать активность отдельных нейронов у мышей из контрольной и тестовой группы, была проведена визуализация участка 300х300 мкм в слоях L2/3 и L4 в момент воздействия звука (чистый тона) 4–64 кГц, 60 дБ (1D). Сначала были определены клетки, которые реагировали на эти стимулы. Клетка, которая хоть как-то отреагировала, классифицировалась как «реагирующая». После воздействия темноты доля реагирующих клеток в L4 не изменилась, а вот в L2/3 на стимул ответило меньше клеток (1E), что говорит об спарсификации (увеличение промежутков между чем-то) корковых ответов в надгранулярных слоях.
Регистрация отдельных единиц* показала, что клетки L4 у мышей из тестовой группы имеют более высокую самопроизвольную и вызванную темнотой частоту нейронов (активность нейронов).
Регистрация отдельных единиц* — метод измерения электрофизиологической реакции отдельного нейрона с применением микроэлектрода.Далее было решено выяснить, присутствуют ли подобные изменения на уровне отдельных клеток в слое L2/3. Чтобы оценить спонтанную активность нейронов (SD), экспрессирующих GCaMP6s мышей, были измерены переходные процессы флуоресценции, предшествующие началу стимула и во время длительной визуализации без какого-либо стимула.
После пребывания в темноте спонтанная активность нейронов увеличилась как в L4, так и в L2/3 (1F).
Снижение тональной чувствительности может быть связано с изменением частотной настройки нейронов слуховой коры мозга. Ранее сделанные записи микроэлектродов показали, что клетки L4 у мышей тестовой группы имеют повышенную частотную избирательность, и, поскольку L2/3 получает входную информацию от L4, такие же изменения могут присутствовать и в L2/3. Следуя этому, были созданы кривые настройки для каждой реагирующей клетки на основе максимального ответа, вызванного воспроизведением звуков (2А).
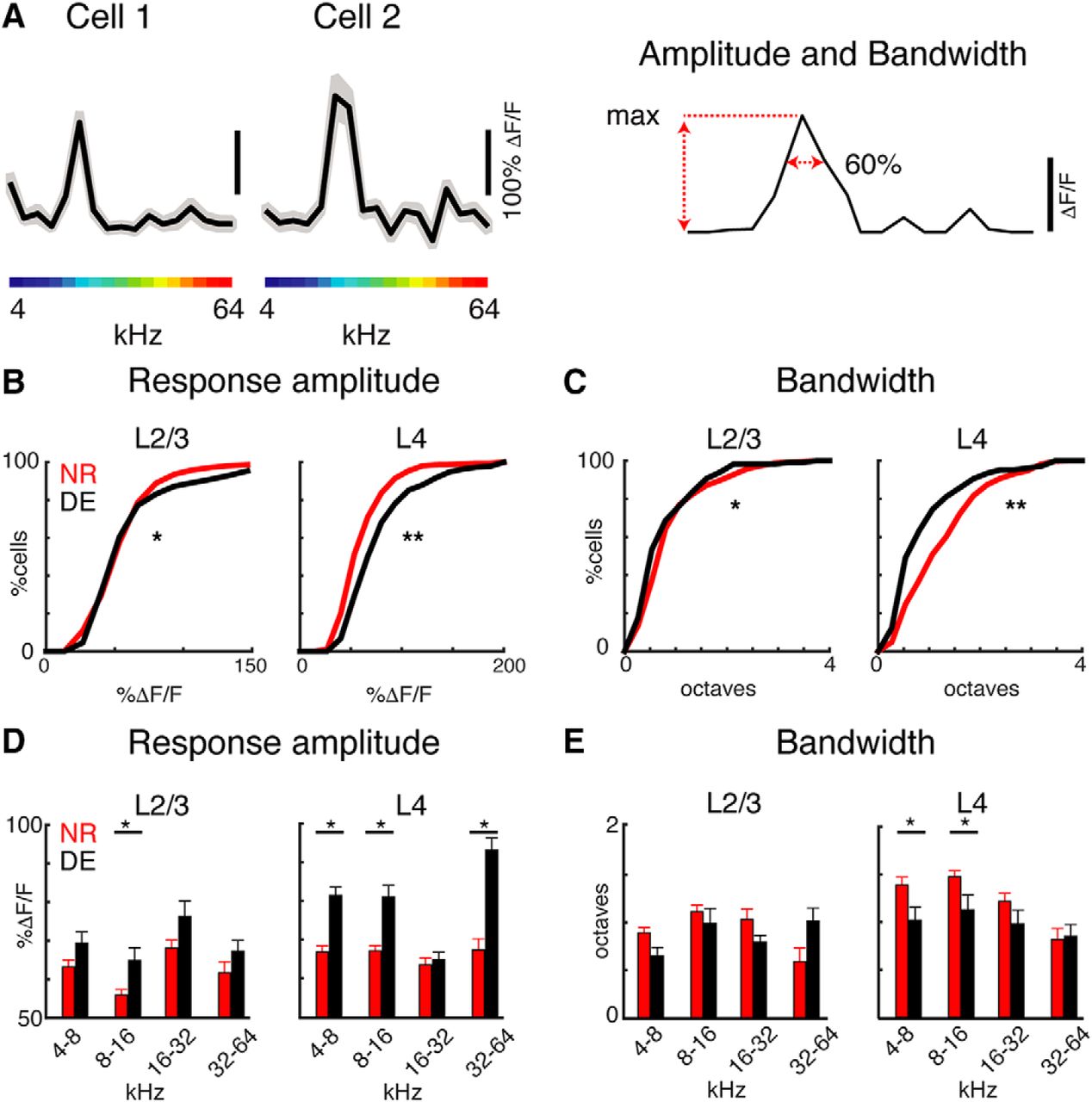
Изображение №2: пребывание в темноте повышает чувствительность и частотную избирательность нейронов как L4, так и L2/3.
Сначала была измерена амплитуда вызванных ответов при лучшей частоте (BF, т.е. пиковой частоте на кривой). Амплитуда ответов после воздействия темноты повысилась и у нейронов L4, и у нейронов L2/3, однако именно в L4 она была больше (2B). Это наблюдение полностью согласуется с электрофизиологическими записями и усилением таламокортикальных афферентов* до L4.
Таламокортикальный резонанс* — явление синхронной осцилляции (периодической одновременной активацией отдельных популяций нейронов) нейронов различных ядер таламуса и связанных с ними областей коры больших полушарий головного мозга.Далее была проведена оценка частотной селективности клеток у мышей из тестовой и контрольной групп. Анализ показал, что полоса пропускания нейронов L4 и L2/3 после воздействия темноты была меньше, чем у животных из контрольной группы (2С).
Афферент* — нейрон, передающий импульсы от рецепторов к головному или спинному мозгу.
Совокупность этих наблюдений говорит о том, что на уровне отдельной клетки изменения после темноты одинаковы как в L4, так и в L2/3, за исключением того, что амплитуды ответа в L4 увеличиваются после темноты, а в L2 / 3 — нет.
Следовательно, после определенного времени воздействие темноты может привести к изменениям вызванных звуком ответов нейронов слуховой коры мозга как в L4, так и в L2/3. Но в слое L4 эти изменения будут все же более значимыми, нежели в L2/3. Хоть после воздействия темноты на звуки реагировало и меньше клеток, те, что отвечали в А1, становились более чувствительными и избирательными к звуку. Другими словами, в темноте нейроны слуховой коры действуют по принципу «качеством, а не количеством», так как число реагирующих клеток снижается, но их активность увеличивается.

Изображение №3: воздействие темноты изменяет распределение частотной селективности в слуховой коре мозга.
Данные наблюдений показывают, что тонально-чувствительные клетки в L4 слуховой коры демонстрируют более высокую амплитуду ответов, а клетки в обоих слоях А1 показали повышенную избирательность к звуку после воздействия темноты. Тем не менее, эти изменения на уровне отдельных клеток не объясняют, почему после воздействия темноты в L2 /3 становится меньше реагирующих нейронов. Нейроны в сенсорных кортикальных слоях могут регулировать свою настройку на основе поведенческих факторов. Более того, ранний сенсорный опыт может изменить площадь слуховой коры, которая реагирует на звуки определенной частоты. Основываясь на этой информации, ученые предположили, что отдельные нейронные популяции изменили свои предпочтения относительно внешних стимулов.
Для проверки этой гипотезы ученые изучили распределение предпочтительных частот для популяций нейронов у мышей из тестовой и контрольной групп.
Мыши из контрольной группы «предпочитали» частоты в диапазоне от 4 до 64 кГц, при этом большинство нейронов предпочитали звуки в диапазоне от 8 до 32 кГц. А вот мыши из тестовой группы активнее реагировали на высокие частоты в диапазоне 32-64 кГц (3B).
В слое L2/3 наблюдался рост числа клеток, более реагирующих на высокие частоты (32–64 кГц), а в слое L4 — на более низкие (4–8 кГц). В обоих слоях наблюдалось снижение восприятия средних частот в диапазоне 8-16 кГц (3А).
Как известно, сенсорные стимулы кодируются не только отдельными нейронами, но и популяциями нейронов. Корреляция активности между нейронами, в свою очередь, способствует процессу кодирования информации. Как в L4, так и в L2/3 соседствующие клетки демонстрируют высокую корреляцию сигналов (SC), которые отражают коррелированную активность, управляемую стимулом. Также существует и парные корреляции (NC), которые представляют независимую от стимула ковариацию между опытами.
Ученые предположили, что улучшение функциональных межслойных и внутриламинарных связей после воздействия темноты может приводить к снижению парной корреляции. Следовательно, NC позволит изучить изменения уровня коррелированной активности между нейронами в L4 и L2/3 после воздействия темноты.
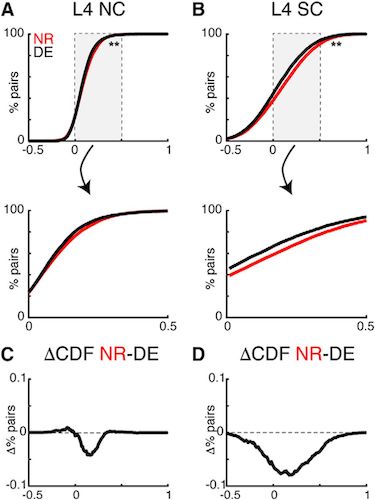
Изображение №4: уменьшение NC в слое L4 за счет воздействия темноты.
Воздействие темноты приводило к снижению NC и SC между клетками слоя L4. Однако, в слое L2/3 NC практически не изменились (5А).
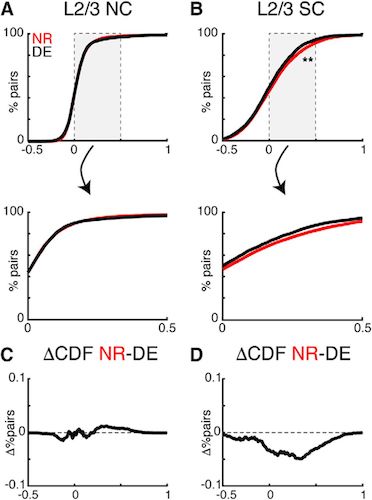
Изображение №5: уменьшение NC в слое L2/3 за счет воздействия темноты.
Темнота приводила к уменьшению SC между нейронами в L2/3 (5B). Из этого следует, что темнота меняет не только настройки отдельных нейронов, но их взаимосвязь друг с другом как в слое L2/3, так и в L4.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
Мыши, пребывавшие в темноте одну неделю, продемонстрировали совершенно иную нейронную активность, нежели мыши, жившие при нормальном освещении. Нейроны слуховой коры мозга мышей из тестовой группы перенастроили сами себя и связи между друг другом для более точного восприятия высоких и низких частот, при этом пожертвовав восприятием средних. Сами ученые не могут пока точно объяснить такую селективность. Они предполагают, что это может быть связано с тем, что именно мыши хотели слышать лучше: свои шаги, звуки от других мышей и т.д.
Важно отметить, что подопытные мыши были здоровы, т.е. их зрение было абсолютно нормальным с самого рождения. Ранее предполагалось, что подобные изменения нейронной активности более вероятны исключительно у тех, кто с самого рождения (или с ранних лет) страдает от какой-либо сенсорной дисфункции. Однако, как оказалось, даже взрослый мозг может измениться, подстраиваясь под условия окружающей среды.
В дальнейшем ученые планируют расширить свое исследование, добавив в него манипуляцию звуками, которые будут слышать мыши. Это позволить более точно определить на какие именно звуки и в какой степени реагируют мыши в темноте.
Данное исследование не только удовлетворяет банальное любопытство ученых, но и может быть крайне полезным для слабослышащих людей. В особенности, этот труд может упростить процесс адаптации слабослышащих к слуховым аппаратам и кохлеарным имплантатам.
Структура, состоящая из миллиардов блоков, способных действовать как отдельно друг от друга, так и совместно, не может быть полностью описана и объяснена за одно исследования. Наш мозг и есть такая система. При этом, с каждым новым исследованием мы узнаем все больше ценной информации касательно нашего важнейшего органа, таинственность которого сравнима с неизведанными просторами Вселенной.
Пятничный офф-топ:
Многие виды животных обладают куда более развитыми органами чувств, чем человек. В данном ролике представлена подборка одних из самых необычных видов, которые могут похвастаться весьма удивительными способностями.
Многие виды животных обладают куда более развитыми органами чувств, чем человек. В данном ролике представлена подборка одних из самых необычных видов, которые могут похвастаться весьма удивительными способностями.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята. :)
Немного рекламы :)
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?