

Изучением представителей флоры и фауны, как правило, занимаются ботаники и биологи. Однако существует множество примеров того, как те или иные существа становились объектом живейшего интереса со стороны физиков, инженеров и математиков. Каждый организм можно назвать системой, которая оказывает определенное влияние на окружающую среду и оказывается под влиянием этой же среды. Все эти взаимодействия тем или иным образом описываются различными химическими реакциями, физическими процессами и математическими формулами. Все становится куда сложнее, когда подобных индивидуальных систем становится много, как в улье пчел или в гнезде муравьев. В таком случае, помимо индивидуального взаимодействия со средой добавляется взаимодействие между индивидами. Группа ученых из Колорадского университета в Боулдере (США) решили изучить рой пчел во время зарождения колонии. Особенность такого роя в том, что множество особей формируют скопление, которое свисает с ветки или другого объекта. Какова динамика такой системы, как ведут себя пчелы, и что помогает им держаться вместе? Ответы на эти вопросы мы найдем в докладе ученых.
Основа исследования
Когда тысячи отдельных насекомых собираются в единую группу, получающийся суперорганизм может выполнять функции, недоступные отдельным особям.
Ярким примером такого суперорганизма является «плот» огненных муравьев, которые они формируют во время затопления.
Другой пример, о котором и пойдет речь, это медоносные пчелы, которые во время миграции колонии скапливаются вокруг королевы, дабы защитить ее.

Медоносные пчелы вне улья.
В этих скоплениях большинство насекомых находятся в основной части конструкции и могут сильно влиять на ее механику. Поскольку невозможно увидеть особей внутри непрозрачных, плотных скоплений, исследования этих скоплений обычно основываются на наблюдениях за особями на поверхности скоплений. В некоторых исследованиях использовались двумерные рентгеновские проекции скоплений, но этот метод не может дать количественную оценку распределения особей во всех трех измерениях.
Авторы рассматриваемого нами сегодня труда решили использовать КТ для визуализации трехмерного расположения медоносных пчел (Apis mellifera) в рое. Это позволило рассчитать, как нагрузка распределяется между пчелами, и почему именно такое распределение задействовано.
Рой медоносных пчел представляет собой группу, состоящую из пчелиной матки и тысяч рабочих, которые висят на ветке дерева (или любом другом объекте) от нескольких часов до нескольких дней, пока рабочие ищут подходящее место для улья. Для нас такой рой является захватывающим зрелищем, но для самих пчел это один из самых опасных этапов жизни. Если в это время матка или слишком много рабочих погибнут, то погибнет и вся колония. Пчелы, буквально хватаются за жизнь, удерживаясь друг за друга, чтобы выдержать вес всего роя и сохранить сплоченность колонии. Следовательно, механическая стабильность этих роев играет важную роль в выживании колонии. И тут возникают важные вопросы — как пчелы распределяются внутри такого скопления, и как вес роя распределяется между ними?
Ранее ученые уже наблюдали за тем, как рой реагирует на изменение среды, т.е. на внешние факторы. К примеру, рой становился площе, если объект крепления (к примеру, ветка) начинал трястись ввиду ветра. Если температура окружающей среды была слишком высока, то особи в рое рассредотачивались, а если становилось холодно, то они наоборот упаковывались плотнее.
Чтобы интерпретировать пространственное распределение пчел, измеренное с помощью рентгеновской компьютерной томографии, ученые представляют рой медоносных пчел в виде суперорганизма, который можно описать с помощью законов масштабирования, аналогичных тем, которые применяются к отдельным организмам в природе.
В биологии был установлен ряд законов масштабирования, связывающих размер особей с их физическими свойствами. Эти законы выводятся из размерного анализа и могут быть проверены экспериментально. Например, скорость метаболизма животных пропорциональна массе тела в соотношении 3 к 4, скорость бега зависит от массы тела в соотношении 1 к 6, а сила тяжелоатлетов пропорциональна массе тела (1 к 3) и т.д. По аналогии с зависимостью между массой тела тяжелоатлета и максимальной поднятой массой скорость, с которой муравьи тянут груз, не является линейно пропорциональной количеству муравьев, тянущих его. Точно так же, как животное — это организм, состоящий из клеток, рой — это сверхорганизм, состоящий из пчел, потому к его физическим свойствам можно применить аналогичные законы масштабирования.
Экспериментальная установка
В наблюдениях приняли участие рои, состоящие из разного числа рабочих (от 3725 до 9700 особей) и королевы. Всего было 11 роев, общее число КТ составило 57. Рой свисал с деревянного диска диаметром 33 см. Средняя масса одной особи колебалась в пределах 0.10-0.13 грамма.
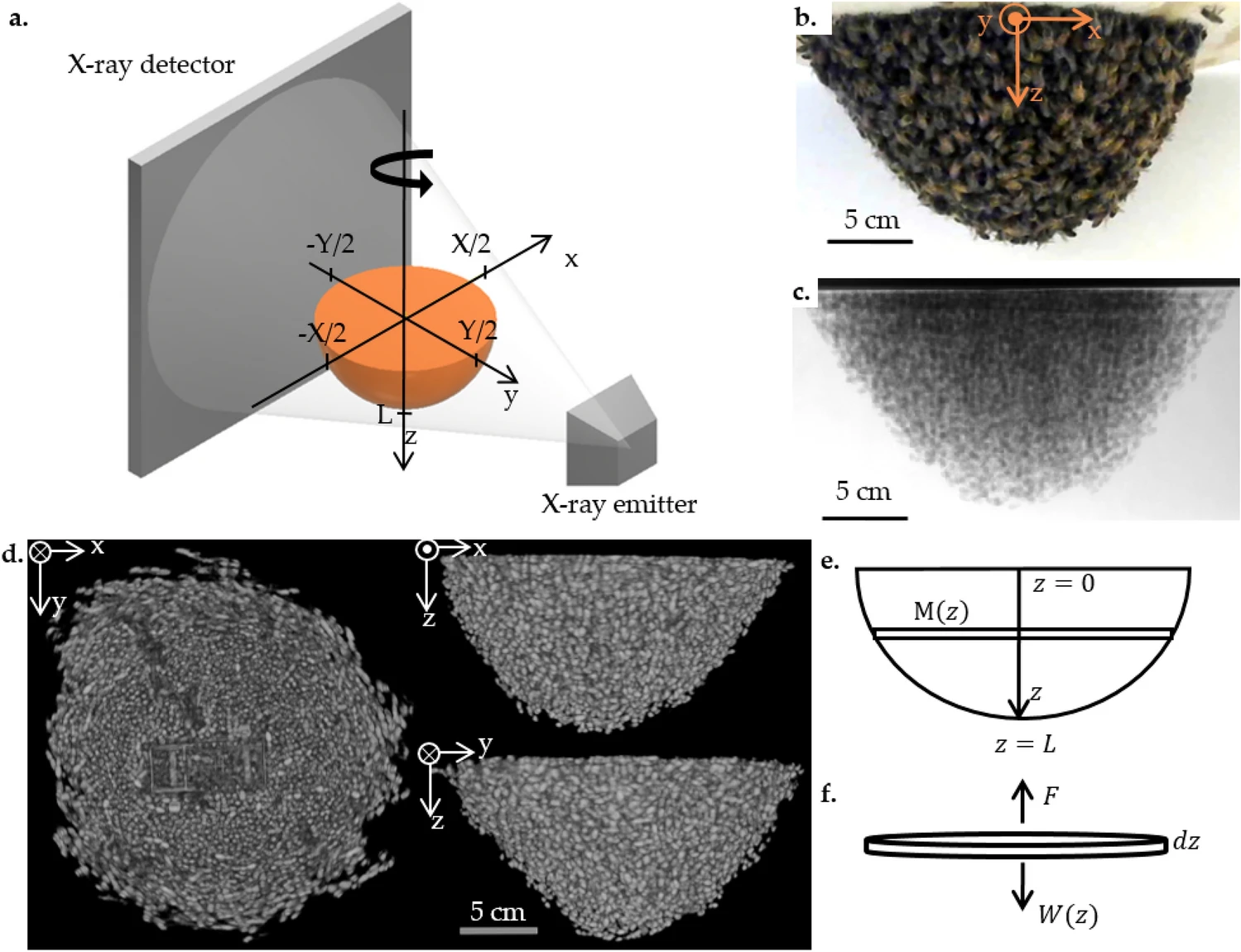
Изображение №1
Каждый рой располагался между рентгеновским излучателем и детектором JPI DynaVue (цифровой рентген и рентгеноскопия в одной системе) на расстоянии в 17.7 ± 0.7 см от детектора. Излучатель и детектор были закреплены на расстоянии 90 см друг от друга (1a).
Ось x соответствовала параллельной плоскости детектора рентгеновского излучения, а ось y — параллельной линии излучатель-детектор, так что поверхность крепления (деревянного диска) и плоскость xy лежали в одной плоскости. Ось z соответствовала направлению силы тяжести с началом в центре диска.
Дополнительно к деревянному диску был подключен шаговый двигатель, способный вращать диск с непрерывной скоростью в 7.2°/с. Во время одного сеанса КТ диск совершал полный оборот за 25 секунд. Во время вращения делались снимки (15 кадров в секунду) с параметрами рентгеновского излучения 90–95 кВ и 20 мА. В качестве наглядной иллюстрации на 1b показан снимок роя, а на 1c — его проекция (видео №1).
Видео №1
Чтобы выполнить 3D-реконструкцию, сначала был найден угол поворота каждой проекции, измерена длина стороны вокселя и центральная координата и вычтено пустое фоновое изображение, чтобы уменьшить шум. Затем проводилась реконструкция каждого набора данных с помощью набора инструментов Matlab TIGRE, используя алгоритм FDK и фильтр Хэмминга. Каждая результирующая реконструкция состояла из трехмерной матрицы значений яркости в каждом вокселе [˜I(x, y, z)].
Были установлены границы реконструкций, чтобы инкапсулировать рой и небольшое количество пустого пространства вокруг него (если только рой не достигает края поля зрения рентгеновского снимка). Ось x ограничена от −X/2 до X/2, а ось y ограничена от −Y/2 до Y/2 (1a). Ось z проходит от 0 до длины роя L (7.7 ≤ L ≤ 12.5 см). X и Y были определены визуально, чтобы инкапсулировать все ненулевые значения яркости КТ (т. е. физическую форму каждого роя). Все воксели роя помечаются, а затем определяется граница поверхности роя. На 1d показано три ракурса реконструкции роя, где видна радиально-симметричная форма роя относительно оси z и очертания отдельных пчел.
Чтобы преобразовать значения яркости I(x, y, z) в меру массы пчел в каждом вокселе, была использована внешняя мера массы всего роя (Mswarm). Предполагая, что яркость I линейно коррелирует с плотностью пчел, следующее соотношение дает расчетную массу на воксель:
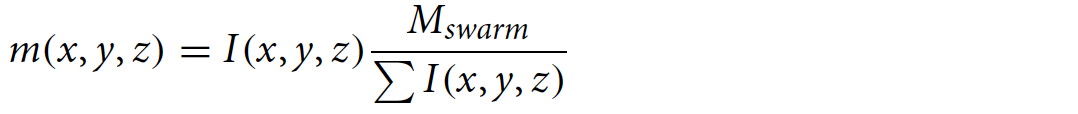
Зная массу каждого вокселя и границу роя, можно определить плотность пчел (z) в каждом слое (xy-срез толщиной в один воксель). Длина стороны каждого вокселя равна s, а NB(z) — это количество вокселей в слое, которые находятся в пределах границы роя. Измерение (z) показывает, насколько плотно расположены пчелы от поверхности крепления до кончика роя:

Также была рассчитана общая площадь, охватывающая каждый слой роя:

Для анализа распределения массы каждого роя использовалась матрица массы в каждом вокселе [m(x, y, z)] путем измерения массы каждого слоя (xy-срез толщиной в один воксель) и веса, поддерживаемого каждым слоем. На схеме 1e показаны три слоя.
Масса пчел на единицу расстояния в каждом слое [M(z)] представляет собой сумму массы каждого вокселя в этом слое, деленную на толщину слоя (s):
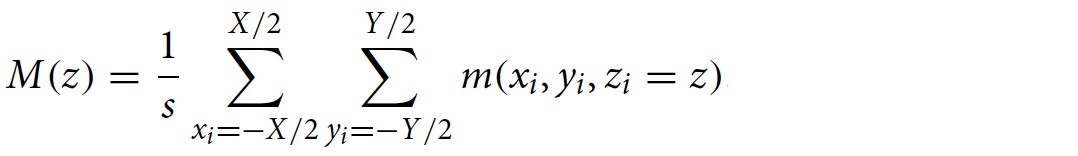
Сила, которую каждый слой роя оказывает вверх, эквивалентна весу, который поддерживает слой (1f). Вес, который должен поддерживать каждый слой, состоящий из веса пчел в этом слое и всех слоев под ним [W(z)] равен:

где g — ускорение свободного падения.
Наконец, измерялась средняя сила, которую каждая пчела поддерживает в слое (Fbee), предполагая, что каждая пчела в слое несет равную долю веса [W(z)]. Поскольку длина ребра вокселя меньше длины пчелы, масса особей, доступных для поддержки пчел под ними, получается с помощью скользящей суммы по вокселям, которые охватывают длину отдельной пчелы (l = 1.5 см). Затем W(z) делится на массу поддерживающих пчел:

Результаты исследования

Изображение №2
В ходе наблюдений было получено 57 КТ одиннадцати роев. Все рои были распределены на три группы по массе (2a): малая, средняя и большая. Затем были проведены реконструкции этих роев под разным углом для демонстрации основания, верхушки и разных сторон роя (видео №2).
Видео №2
Первый вопрос, на который хотели получить ответ ученые, это — как пчелы распределяются по рою, чтобы выдержать его вес? Чтобы ответить на этот вопрос, необходимо было вычислить массу пчел в каждом слое роя [M(z)] и вес, поддерживаемый этими слоями [W(z)].
На 2b построен график зависимости W(z)от M(z) в логарифмическом масштабе для роев весом 447–970 г, содержащих от 3725 до 9700 особей. Несмотря на разные размеры роя, все кривые W(z) и M(z) падают на универсальную линию. Это позволяет предположить, что рои имеют одинаковую фундаментальную структуру в малых и больших масштабах. Чтобы дополнительно охарактеризовать данный закон масштабирования, ученые применили линейное приближение к каждой кривой. Наклон этой подгонки представляет собой показатель степени a в степенном законе, связывающем M(z) с W(z):

где a = 1.53 ± 0.12 и константа C = 250 ± 110. Затем а для каждой массы роя был нанесен на 2с. Это показало, что тенденции в зависимости от массы роя нет. Это еще раз подтверждает, что распределение веса не зависит от массы роя.
На 2b показано, что слои с наибольшим M(z) поддерживают наибольший вес роя W(z). Соответственно, три реконструкции на 2а и график площади каждого слоя на 2e показывают, что площадь слоев роя вблизи поверхности крепления к деревянному диску больше, чем площадь слоев вблизи вершины роя.
Чтобы количественно оценить влияние плотности упаковки на массу слоя, был построен график плотности каждого слоя в зависимости от координаты z для всех 57 реконструкций (2d). Пчелы были наиболее плотно собраны в верхней части роя, при этом плотность уменьшалась по направлению к кончику роя. Это подтверждает, что большинство пчел находятся у основания роя.
Чтобы количественно оценить расположение слоев роя от поверхности прикрепления до кончика роя, что приводит к степенному закону, была построена зависимость M(z) от z (3a). M(z) высока вблизи поверхности прикрепления (z ≤ 2 см), а затем уменьшается с z к вершине роя. Это видно в рентгеновских проекциях роя, например, на 1c. Более темный оттенок серого представляет большее количество пчел в горизонтальном слое между рентгеновским излучателем и детектором излучения, а белый цвет означает отсутствие пчел. В этой проекции имеется широкий слой более темного серого цвета, представляющий большую массу пчел на горизонтальный слой вблизи поверхности прикрепления, за которым следуют более узкие слои светло-серого цвета, представляющие меньшее количество пчел на горизонтальный слой по направлению к кончику роя. Большая масса пчел в самом верхнем ярусе создает устойчивую опорную конструкцию для основной массы роя, распределяя нагрузку между пчелами.
Зная эту информацию, ученые задались следующим вопросом — как это расположение влияет на силы, испытываемые отдельными пчелами в рое? Если вес, поддерживаемый отдельной пчелой, близок к ее пределу, эта пчела с большей вероятностью разорвет свою связь, когда рой будет механически возмущен.
Чтобы выяснить, какой вес выдерживают отдельные пчелы, мы вычисляем среднюю силу, поддерживаемую пчелой в слое [Fbee(z)]. В результате был построен график зависимости Fbee от z (3b), по которому видно, что каждая пчела поддерживает максимум 3.8 собственного веса. Вес, поддерживаемый одной пчелой, уменьшается от места крепления (основания роя) к кончику роя. Ранее ученые провели опыты, в которых определили, что максимум, который одна плеча может поддержать составляет 35 ± 14 собственного веса. Следовательно, пчелы в рое не достигают своего предела возможностей.
Вышеописанные экспериментальные данные показывают закон масштабирования между массой слоя по вертикальной координате, M(z), и весом, который он поддерживает: W(z) ∼ M(z)a с a ≈ 1.5.
Ученые высказывают предположение, что рой находится в квазиравновесии (форма не меняется, хотя отдельные пчелы могут двигаться), что все пчелы в каждом слое в равной степени способствуют поддержанию веса пчел под этим слоем, что толщина слоя очень велика, и что рой радиально симметричен относительно оси z.
Потому была использована цилиндрическая система координат с вертикальной координатой z (1e) для рассмотрения слоев роя вдоль оси z толщиной dz. Также ученые отмечают, что переменные, помеченные тильдой [например, ˜W(z)] представляют собой аналитически полученные выражения, а переменные без тильды [W(z)] представляют значения, определенные с помощью степенной зависимости от экспериментальных данных.
Анализ данных был начат с применения принципа баланса сил к каждому слою роя. Как показано на диаграмме свободного тела на 1e, сила, с которой каждый слой пчел должен держать слой над ним, равна весу этого слоя и всех слоев под ним: ˜F = ˜W(z). ˜W(z) можно выразить с помощью уравнения баланса сил:

где масса пчел на ярус равна ˜M(z), длина роя равна L, а g — гравитационная постоянная.
Учитывая экспериментальные наблюдения, где масса слоев у основания самая высокая, а масса слоев на вершине роя самая низкая (3a), была смоделирована ˜M(z) как монотонно убывающую функцию z. Чтобы единицы измерения были согласованными, координата z была нормализована по длине роя:

А показатель степени в выражении для ˜M(z) будет равен:

Тогда вес, поддерживаемый каждым слоем, равен:

Прочность слоя [˜S(z)] или максимальный вес, который он может выдержать, может быть больше или равен весу роя [˜S(z) ≥ ˜W(z)]. Если бы вес пчел под слоем превышал его силу ˜S(z), слой не смог бы выдержать вес этих пчел, и рой распался бы. Сила пропорциональна массе, которая пропорциональна объему или кубу длины, поэтому прочность слоя зависит от куба длины, ˜S(z) ∝ L3. Масса каждого слоя пропорциональна площади или квадрату длины, поэтому ˜M(z) ∝ L2. Следовательно, ⍺ должно быть равно 1.5, чтобы ˜S(z) ∼ ˜M(z) было правильным с точки зрения размерности.
Оценка ˜W(z)/˜S(z) дает меру того, какую часть своей максимальной силы каждый слой использует для удержания остальной части роя:

Среднее количество пчел, которое поддерживает одна особь в слое роя, ˜Fbee(z), равно массе пчел, поддерживаемых слоем, деленному на сумму массы пчел в слое, толщина которого равна длине одной особи (l ≈ 1.5):

После интегрирования было получено выражение для ˜Fbee(z):
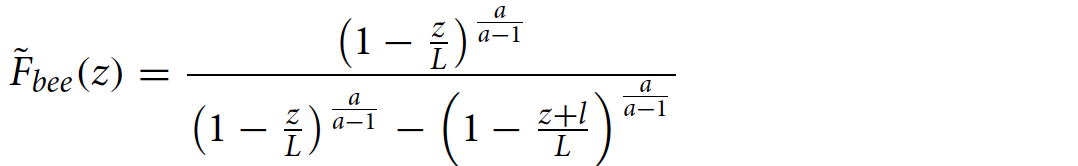
Далее ученые рассмотрели влияние a на массу каждого слоя, долю его максимальной силы, которую он использует, и среднюю силу на одну пчелу.

Изображение №3
Чтобы визуализировать влияние a на распределение пчел, был построен график массы на слой длиной 12.5 см из роя в 1000 грамм, ˜M(z) в зависимости от z/L, с a = 1.5, 1.01, 1000 и -0.2 (3c) и соответствующего среднего усилия на пчелу (Fbee(z) против z/L на (3d).
Если a ≈ ⍺, как это показывают экспериментальные данные, слои с большей массой вблизи поверхности прикрепления поддерживают менее массивные слои под собой, как это показано сплошной черной линией на 3c. Соответственно, на 3d видно, что ˜Fbee(z = 0) ≈ 3 в верхней части роя и уменьшается к вершине. Сила каждого слоя и вес, который он поддерживает, пропорциональны друг другу (˜W(z)/˜S(z) ∼ 1/3). Это означает, что доля максимальной силы, используемая слоем, одинакова для всех z.
Видео №3
Если 1 < a < ⍺, рой близится к одному массивному слою пчел, как показано фиолетовой пунктирной линией на 3c. Анализ прочности такого роя показывает крайне малые значения: ˜W(z)/˜S(z) → 0(1 − z/L )−∞. Сила, поддерживаемая каждой пчелой на 3d, показывает ˜Fbee(z) = 1 для всего роя. Это означает, что каждая пчела поддерживает только свой собственный вес. Эта конфигурация потребует упаковки большого количества пчел либо в один очень плотный, либо в один очень широкий слой. Рой с одним очень плотным слоем наверху сжал бы всех пчел; рой с одним очень широким слоем потребует большой площади поверхности, что подвергнет рой опасности со стороны хищников и изменений погоды.
При значениях a > ⍺ (при a→∞) все слои роя имеют одинаковую массу (красная линия на 3c). Сила на пчелу на 3d показывает ˜Fbee(z = 0) ≈ 8 на вершине роя, что в 2.5 раза больше, чем в конфигурации a = ⍺. В этой конфигурации верхние слои используют более высокий процент доступной прочности, чем нижние слои. Таким образом, для больших роев пчелы, поддерживающие рой, будут подвергаться большему напряжению, и рой с большей вероятностью разрушится под внешним возмущением.
Видео №4
Наконец, a < 0 предполагает, что верхние слои роя имеют меньшую массу, чем нижние слои (оранжевая линия на 3c). Это нереалистичный диапазон значений a, но они были включены в анализ как демонстрацию потенциального распределения массы, когда самые большие слои находятся на дне роя. Эта конфигурация создаст еще большую нагрузку на слои пчел в верхней части роя, поскольку меньшие слои вблизи поверхности прикрепления имеют меньшую максимальную силу. Соответственно, средняя сила, приходящаяся на одну пчелу (3d), превышает максимум ее возможностей, потому рой не может поддерживать себя в такой конфигурации.
Из вышеописанного анализа следует, что конфигурация роя с a ≈ 1.5 использует полную силу каждого слоя и создает меньшую нагрузку на пчел, чем большинство других значений a, и позволяет избежать распределения веса, которое может подвергнуть большое количество пчел внешней опасности.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
На первый взгляд может показаться, что, к примеру, работа двигателя самолета намного сложнее процесса роста дерева. В двигателе множество деталей, каждая из которых должна быть расположена в определенном месте и выполнять определенную функцию, чтобы сам двигатель мог исправно работать. Но и в обычном дереве протекает множество сложных процессов, которые обеспечивают его рост. Этот незамысловатый пример лишь показывает, что природа куда сложнее, чем мы о ней думаем.
Авторы рассмотренного нами сегодня труда подтвердили это. Ученые решили просканировать с помощью КТ рой пчел, прикрепленный к деревянному диску. Рой во главе с королевой собирается в многотысячное формирование и крепится к веткам деревьев или другим объектам, пока часть рабочих занята поиском подходящего места для создания нового улья. Ученые называют это скопление пчел суперорганизмом. А каждую пчелку можно по аналогии назвать клеткой этого организма. Следовательно, чтобы организм не рассыпался, все клетки должны держаться друг друга, поддерживая целостность подвесного роя.
Мнение, что структура роя является случайно, как показало исследование, является ошибочным. Пчелы создают куполообразную структуру, следуя удивительно сложным математическим правилам, или тому, что исследователи называют «законом масштабирования». Суть в том, что пчелам необходимо расположиться и держаться друг за друга так, чтобы не превышать нагрузочный максимум друг друга и своего слоя в целом. У каждого слоя в рое этот показатель свой, так как он должен держать не только себя, но и все нижевисящие слои.
Подобные примеры реализации закона масштабирования не редкость в природе, но они пока малоизученны. Потому ученые намерены продолжить свой труд, так как его результаты могут стать крайне полезны для инженеров, проектирующих более устойчивые здания, или для робототехников, создающих роящихся микроботов.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Комментарии (7)

excoder
04.11.2022 16:15Отличная статья! Вспоминается ещё одна недавняя про самоорганизацию:
https://habr.com/ru/post/694156/

vip100
05.11.2022 14:24Может быть это просто "ошибка выжившего" ? Пчелы по всякому организуют рой, но неудачные быстро рассыпаются и ученые их не видят ? Возможно они не рассыпаются полностью, и те кто отвалился просто переползают в другое место. Для внешнего наблюдателя это даже может быть незаметно.

dengus
05.11.2022 23:00По факту пчёлы висят кучей не только во время роения, а по большей части, например, в жаркий период под вечер частенько выкручиваются на прилетках улья для регулирования температуры внутри улья. Кстати очень необычно такую висящую кучу пчел рукой трогать и полную горсть набирать - как они лапками сцепляются друг с другом.

HOLAOLA
06.11.2022 11:31Эммм...так они не кусаются в этот момент ?
dengus
07.11.2022 11:01Для пчел не совсем корректно слово кусаются, они жалят. В конкретном случае, когда происходит выкучивание, то если их аккуратно трогать и даже всю кучу загребать ладонью, то может ужалить несколько пчел, а могут и не ужалить вообще. Хотя по опыту скажу, что работая на пасеке, бывает ужалят в руки сразу десяток-два пчел и не больно, а бывает одна ужалит и очень больно. В любом случае боль через минуту проходит. А на яд пчелиный уже иммунитет есть )) Вообще, если пчелам не мешать - не надевать темное, не шуметь и не делать резких движений, то на человека они не обращают внимания. Но не факт )) Конечно от породы пчел агрессивность тоже зависит.

KabirK
07.11.2022 10:17законы масштабирования — это всё-таки чисто эмпирические зависимости, а структура роя (стаи, муравейника, города…) — это динамическая система. в области complexity много интересных и красивых исследований, много математических примеров того, как простые зависимости приводят к сложным структурам, которые в пределе становятся неотличимы от случайных (а до этого предела демонстрируют то, от чего захватывает дух, — мурмурацию, например).
тут следовало бы говорить о роевом интеллекте, суть которого в том, что пчёлы (птицы, рыбы, муравьи, люди) не следуют «удивительно сложным математическим правилам», а следуют очень простым правилам, которые в итоге дают очень сложную картину. на мой взгляд, несколько странный акцент у этого исследования, притом что один из авторов — из Santa Fe Institute, а это мировой лидер в области complexity.
InBioReactor
Классная статья. Прочитал с удовольствием