
Введение
Для эукариотических геномов характерно наличие большого количества интронов, микро- и минисателлитов, диспергированных элементов, «реликтовых ретровирусов» и прочих нуклеотидных последовательностей, суммарная длина которых более чем на порядок превосходит длину генных экзонов, определяющих первичную структуру белков. Обычно считают, что избыточная ДНК является реликтом эволюции и только засоряет клеточное ядро. В лучшем случае ей отводится роль радиопротектора, защищающего гены от повреждений /1/.
Такие объяснения достаточно правдоподобны, но недостаточно убедительны. Трудно убедить себя в том, что наш геном является мусорным ящиком, набитым эволюционными отбросами или даже вредными для организма эгоистичными нуклеотидными последовательностями. Возможная радиопротекторная роль этого “мусора” служит слишком слабым утешением.
Особенно контрастно избыточность генома человека проявилась после завершения его секвенирования. Выяснилось, что в гаплоидном наборе хромосом содержится меньше 30 тысяч генов, и на каждый ген приходится примерно 100 тысяч нуклеотидов. Это значит, что в кодировании белков принимает участие менее 2% хромосомной ДНК.
Парадоксальность избыточности генома и непостижимость молекулярных основ человеческого разума являются самыми интригующими загадками современной биологии. А что, если эти две загадки имеют общую разгадку?
Компьютерные аналогии
Попытки понять принципы работы человеческого мозга, как правило, основываются на поиске аналогий между строением центральной нервной системы и устройством компьютера. Нервам при этом отводится роль проводников электрических импульсов, а клетки головного мозга приравниваются к ячейкам хранения и обработки цифровой информации. Но в основе способности мозга воспринимать и перерабатывать информацию могут лежать и иные принципы. Например, уже почти забыты аналоговые компьютеры, позволявшие моделировать некоторые технологические и даже биологические процессы с помощью сопротивлений, конденсаторов, транзисторов и прочих элементов электрических схем. Возможны и другие способы обработки как цифровой, так и аналоговой информации.
В последние годы повышенное внимание уделяется теоретической и экспериментальной проработке идеи квантового компьютера, оперирующего не с битами, а с кубитами, т.е. с квантовыми битами, носителями которых служат фотоны, электроны, атомы или возбуждённые молекулы. Содержащаяся в кубитах информация обрабатывается параллельно, что требует использования специальных алгоритмов, но реализация данного подхода позволит создавать суперкомпьютеры микроскопических (молекулярных) размеров, способные практически мгновенно выполнять сложнейшие вычисления /2/.
Разумеется, мозг вряд ли содержит квантовые логические элементы атомного или фотонного типа, но в основе мышления может лежать взаимодействие между собой сложных молекул, приводящее к изменению их химической структуры или конформации. Молекулярные логические операции должны иметь много общего с квантовыми вычислениями и с особенностями работы квантовых компьютеров, к которым можно отнести параллелизм обработки информации, нанометровые размеры логических элементов, а также высокую эффективность вычислений. Для некоторых алгоритмов эффективность квантовых компьютеров на порядки превосходит производительность современных цифровых суперкомпьютеров, причём это достижимо при наличии всего нескольких десятков или сотен логических элементов /3/. Следовательно, эффективность обработки информации не связана с размерами вычислительного устройства.
Центральный биопроцессор
При современном уровне развития молекулярной биологии очень трудно представить себе, что в мозгу человека смогли остаться незамеченными какие-то “умные молекулы”, запоминающие и перерабатывающие поступающую извне информацию. Если мышление действительно основано на молекулярных элементах, то их химическую природу предсказать не так уж и сложно, так как словосочетание “информационные молекулы” уже давно и прочно ассоциируется с ДНК и РНК. Именно нуклеиновые кислоты способны с высокой специфичностью взаимодействовать между собой, реплицироваться, транскрибироваться, приобретать сложную вторичную и третичную структуру, изменять конформацию, фрагментироваться и даже редактироваться.
Условия для взаимодействия информационных молекул имеются только в клеточном ядре. В нем сосредоточена вся хромосомная ДНК и значительная часть РНК. Это означает, что именно ядро способно служить центральным биопроцессором.
Цитоплазма отделена от ядра мембраной, которая с высокой избирательностью транспортирует участвующие в экспрессии генов молекулы РНК, т.е. те самые молекулы, назначение которых вполне понятно. Следовательно, цитоплазма не содержит (или почти не содержит) информационных молекул, кодируемых избыточной частью генома, и может претендовать только на роль периферийного устройства, соединённого с биопроцессором.
Если подобное предположение справедливо, то многоклеточный эукариотический организм следует считать мультипроцессорным устройством, состоящим из множества автономных ядерных биопроцессоров. Информация от ядра к цитоплазме передаётся в форме мРНК, кодирующей белки, но для межклеточного (межъядерного) обмена информацией должны использоваться какие-то вторичные химические или физические сигналы, поскольку ограничивающие цитоплазму клеточные мембраны, как правило, непроницаемы для нуклеиновых кислот.
Молекулярный биокомпьютер
Идея использования нуклеиновых кислот в качестве молекулярных элементов вычислительных устройств не нова /4, 5/. Существует даже теория волнового генома, приписывающая ДНК способность воспринимать, обрабатывать и излучать информацию в виде модулированного электромагнитного излучения /6/. К сожалению, подобные идеи и теории не позволяют понять сущность элементарных логических операций, на которых может быть основано ядерное мышление. Кроме того, эти операции могут оказаться совсем не элементарными, так как алгоритмы работы простейшего квантового компьютера с большим трудом воспринимаются немногими специалистами, а алгоритмы работы молекулярного биокомпьютера могут оказаться на несколько порядков сложнее.
Взаимодействие информационных молекул между собой происходит параллельно, и описать последовательность событий практически невозможно по причине отсутствия самой последовательности. Представьте себе, что Вы прочли этот текст не по словам, а восприняли его целиком. Затем, не делая отдельных умозаключений, измените своё представление об окружающем мире. Возможно, именно так работает ядерный биопроцессор, у которого результатом обработки информации является изменение состояния системы информационных молекул.
Структуральная лингвистика
Последовательности нуклеотидов, обеспечивающие информационные функции, должны подчиняться каким-то “грамматическим” правилам. По-видимому, существуют и правила “орфографии”, определяемые закономерностями формирования вторичной и третичной структуры нуклеиновых кислот, а также соотношением структуры молекул с их информационными свойствами. Изучение подобных правил может стать основной задачей структуральной лингвистики /7/.
В эукариотических геномах содержится до 50 % сателлитной ДНК, состоящей из блоков тандемно повторяющихся нуклеотидных последовательностей. Различают микросателлитную (до 4 п.о. в повторе) и минисателлитную (более 4 п.о.) ДНК. Блоки комплементарных нуклеотидных повторов могут взаимодействовать между собой и участвовать в межмолекулярных взаимодействиях, необходимых для биопроцессорной обработки информации, и/или в регуляции экспрессии генов /8/.
К другому типу повторов относятся последовательности, подразделяемые на SINE (короткие) и LINE (длинные) диспергированные элементы. Длина SINE-элементов находится в пределах 400 п.о., а у LINE- элементов достигает 7 000 п.о. В геноме человека содержится около 1 млн. Alu-повторов, относящихся к SINE-элементам, и более 100 тысяч длинных MaLR-повторов /1/.
Общими чертами диспергированных элементов являются их транскрибируемость и способность к транспозиции. Эти особенности могут определяться участием таких нуклеиновых кислот в работе ядерного биопроцессора. Транскрибируемость, например, может быть связана с необходимостью перевода двунитевой ДНК в однонитевую РНК, способную к межмолекулярным взаимодействиям. Транспозиция способна перемещать диспергированные элементы и изменять их взаиморасположение, приводя к функциональной адаптации структуры биопроцессорной ДНК.
Наиболее загадочной особенностью структуры эукариотических генов является их мозаичная интрон-экзонная структура. Гены без интронных вставок являются у эукариот довольно редким исключением. Экзоны, как правило, кодируют транслируемые последовательности, а вот назначение интронов совершенно непонятно. Наиболее привлекательная гипотеза объясняет мозаичность генов существованием сложной системы регуляции их экспрессии на уровне сплайсинга, причём в подобной регуляции принимает участие взаимодействие интронов с ещё не изученными регуляторными последовательностями /9/.
Средняя длина экзонов составляет 137 п.о., тогда как длина интронов может превышать 100 000 п.о. /1/. Считается, что в результате сплайсинга интроны удаляются из мРНК, но в большинстве случаев правильнее рассматривать этот процесс как контролируемое удаление экзонов из мозаичной молекулы, основная часть которой представлена интронами.
После сплайсинга мРНК переходит в цитоплазму, а многочисленные петлевидные молекулы интронной РНК остаются в ядре. Подобные молекулы способны нести информацию о количестве и характере РНК, поступившей в цитоплазму, и/или непосредственно регулировать созревание пре-мРНК через сложную сеть специфических межмолекулярных взаимодействий. Нельзя исключить и вероятность существования глобальной мультимолекулярной многофункциональной биопроцессорной сети, в которой интроны функционируют в качестве регуляторов экспрессии генов, причём подобная регуляция осуществляется как через сплайсинг, так и через многочисленные энхансеры (усилители) и сайленсеры (ингибиторы) транскрипции.
Синтезируемая в ядре РНК объединяется в гетерогенные ядерные рибонуклеопротеиновые комплексы (гяРНП), ассоциированные с компонентами аппарата сплайсинга и ядерным матриксом. Для комплексов гяРНП характерно диффузное распределение в цитоплазме, но часть из них может концентрироваться в окрестностях спеклов (см. далее) и даже перемещаться в цитоплазму и снова возвращаться в ядро.
Некоторые факты указывают на то, что перемещение РНК от гена к ядерной мембране не является следствием простой диффузии. Да и расположение генов, в соответствии с моделью ядерной фиксации (gene gating model), может быть функционально связано с определёнными участками (и порами) ядерной мембраны /1/.
Архитектоника ядра
Ядро окружено сложной оболочкой, к внутренней стороне которой прилегает электронно-плотный слой ядерной ламины. Пронизанная белковым скэффолдом (скелетом или ядерным матриксом) нуклеоплазма содержит хроматин — упорядоченные структуры, содержащие ДНК, а также гистоны и негистоновые белки.
В зависимости от степени компактизации ДНК различают эухроматин, в котором экспрессируется основная часть генов, и гетерохроматин, обладающий пониженной транскрипционной активностью. Гетерохроматин, в свою очередь, подразделяется на конститутивный, который часто состоит из повторяющихся последовательностей ДНК, и факультативный, формирующийся на определённых стадиях жизненного цикла организма и обычно присутствующий лишь в одной хромосоме из пары гомологов. Структурно отличающиеся участки хроматина могут быть фланкированы инсуляторами – пограничными последовательностями нуклеотидов, принимающими участие в активации транскрипции генов.
У эукариот выделяют три уровня структурной организации хроматина: 1) нуклеосомная фибрилла; 2) соленоид (нуклеомер); 3) петельно-доменная структура, включающая хромомеры /1/. Хромомеры формируются в результате специфического взаимодействия ядерного матрикса с MAR/SAR-последовательностями ДНК (Matrix Associated Region / Scaffold Associated Region) /8/. Топологические домены, выделяемые MAR/SAR-последовательностями, могут изменяться в процессе индивидуального развития организма, что сопровождается изменением характера экспрессии больших блоков генов в дифференцирующихся клетках. Кроме того, эти последовательности участвуют в формировании оси хроматиды, образующей вместе с петлями хроматина тело метафазной хромосомы /1/.
Распределение участков гетеро- и эухроматина в интерфазных хромосомах упорядочено и, по-видимому, может служить видовым признаком. Теломерные участки хромосом находятся вблизи оболочки, тогда как активно экспрессирующиеся гены локализуются преимущественно во внутренних частях интерфазных ядер. Показано, что отдельные хромосомы в ядре не смешиваются и занимают дискретные зоны, на периферии которых находятся компоненты аппарата сплайсинга и треки синтезируемой РНК /1/.
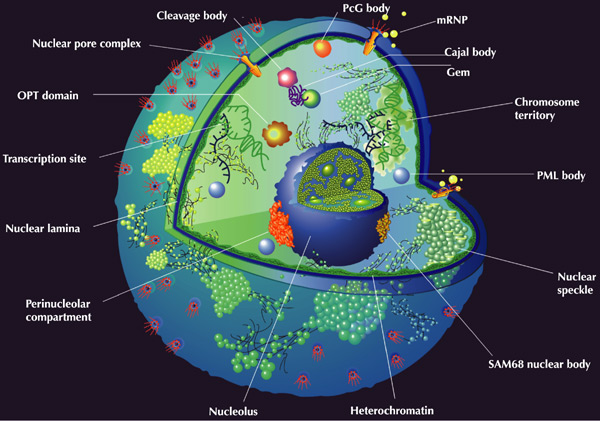
Наиболее крупным нехромосомным компартментом ядра является ядрышко – нуклеопротеиновый комплекс, в котором различают три основные высокоупорядоченные зоны: фибриллярный центр, окружающую его фибриллярную зону и внешнюю гранулярную область. Считается, что наблюдаемые изменения структуры ядрышка определяются его функцией внутриядерного микрокомпартмента биогенеза рибосом. В последнее время появились данные, указывающие на участие ядрышка в регуляции клеточного цикла /1/.
Морфологически в нуклеоплазме различают клубки переплетённых нитей, которые часто ассоциированы с периферией ядрышка (свёрнутые тельца), плотные фибриллярные кольца (ядерные тельца PML), ядерные домены WT1 и тельца Барра (компактный гетерохроматин неактивной X-хромосомы). На границах доменов конденсированного хроматина располагаются перихроматиновые фибриллы — рибонуклеопротеиновые комплексы диаметром 3…20 нм. Здесь же обнаруживают компоненты аппарата сплайсинга, удаляющего интроны из предшественников мРНК. Кроме того, синтез РНК тесно ассоциирован с дискретными межхроматиновыми гранулами, называемыми спеклами, причём каждое ядро содержит от 20 до 50 спеклов /1/.
К сожалению, изучение ядерной архитектоники сопряжено с большими методическими трудностями, но накопленной информации вполне достаточно, чтобы констатировать чрезвычайно высокую сложность структурной организации клеточного ядра. Это не может служить доказательством существования ядерных биопроцессоров, в которых информационные потоки обрабатываются молекулярными логическими элементами, но позволяет не отвергать подобную возможность a priori.
Разумная изменчивость
Если в процессе эволюции возникла молекулярная система, обладающая элементами сознания, то её главной задачей должно было стать самосовершенствование. Это могло привести к появлению механизмов редактирования нуклеотидных последовательностей, изменяющих кодирующий потенциал мРНК или целенаправленно воздействующих на основания геномной ДНК.
В последние годы обнаружено несколько механизмов редактирования РНК на посттранскрипционном уровне. Показано, например, что у митохондрий слизневиков во множественные участки РНК встраиваются остатки цитозина. В митохондриях трипаносом в редактировании РНК, вероятно, принимают участие сложные нуклеопротеидные комплексы – эдитосомы /1/.
В митохондриальных и ядерных транскриптах позвоночных животных при полиаденилировании мРНК появляются бессмысленные кодоны, приводящие к преждевременной терминации трансляции и к синтезу новых полипептидных цепей. В митохондриях высших растений в результате ферментативного дезаминирования некоторые остатки C заменяются на U. Подобный механизм редактирования мРНК обнаружен и в хлоропластах /1/.
Изучение посттранскрипционного редактирования мРНК ещё только начинается, но вполне вероятно, что подобный феномен, встречающийся и у прокариотических микроорганизмов, для эукариот является правилом, а не исключением. Гораздо более проблематичным является вопрос о наличии у эукариот молекулярных механизмов коррекции геномной ДНК, способных влиять на эволюцию животных и растений.
Существование направленной (адаптивной) генетической изменчивости отрицается современной наукой, поэтому обнаружение статистически достоверного повышения частоты появления Lac+ ревертантов кишечной палочки на среде с лактозой /9/ чуть не стало “ересью”, способной оживить старый спор неоламаркистов с неодарвинистами. Но всё благополучно завершилось появлением нескольких гипотез, объясняющих данный феномен на основании постулата о ненаправленности изменчивости, доказанного ещё в середине прошлого века при помощи экспериментов всё с той же кишечной палочкой.
Ненаправленность изменчивости, являющаяся одной из основных аксиом синтетической теории эволюции, имеет много оппонентов /12, 13/. Их возражения не противоречат доказательствам, полученным в экспериментах с прокариотами, так как основаны преимущественно на необъяснимых с позиций неодарвинизма закономерностях эволюции эукариот. К сожалению, одной только констатации подобных фактов недостаточно для доказательства разумности генетической изменчивости эукариот. Современная наука требует, чтобы подобные доказательства опирались на достижения молекулярной генетики. Для этого необходимо рассмотреть механизмы модификации ДНК, которые могут использоваться эукариотами для самосовершенствования.
Простейшим вариантом генетической адаптации является обратимая дупликация генов, характерная как для бактерий, так и для эукариот. Дупликация генов, детерминирующих синтез рибосомной РНК, контролируется на молекулярном уровне, но этот вид адаптивной изменчивости можно причислить к обычным механизмам регуляции экспрессии генов.
Более сложные адаптивные модификации генома могут быть связаны с метилированием ДНК. Содержание метилцитозина в ДНК эукариот достигает 7% /11/. Эукариотическая (цитозин-5)-ДНК-метилтрансфераза действует преимущественно на двунитевую ДНК, одна из нитей которой уже содержит метилированный динуклеотид СpG. Такая специфичность фермента обеспечивает наследуемость признаков, определяемых заменой цитозина на метилцитозин. Метилирование ДНК является одним из основных факторов тонкой регуляции экспрессии генов и, вполне вероятно, способно участвовать в эпигенетической адаптации, ускоряющей эволюцию эукариот. Косвенным указанием на участие метилирования не только в микроэволюции, но и в формировании новых видов, может оказаться до неприличия высокое сходство секвенированных генов человека и шимпанзе.
Пока нет никаких доказательств участия метилирования ДНК в адаптивной изменчивости эукариот, но участие его в эпигенетическом наследовании некоторых признаков уже не вызывает особых сомнений. Метилирование вполне может лежать в основе некоторых проявлений адаптивной эволюции эукариот. Вопрос только в том, насколько подобная адаптация управляема, т.е. способен ли геном модифицировать сам себя, причём не методом проб и ошибок, а вполне целенаправленно.
Один из уже известных механизмов целенаправленной адаптации эукариотических клеток является диминуция хроматина, т.е. запрограммированная потеря значительной части хромосомной ДНК при формировании некоторых многоклеточных организмов /14/. Рекорд по доле элиминируемого генома пока принадлежит циклопу Cyclops kolensis. В ядрах соматических клеток этого циклопа сохраняется всего лишь 6% генома стволовых клеток /15/. Менее выраженные генетические перестройки и делеции участков хромосом соматических клеток являются скорее правилом, чем исключением. Главной особенностью подобных генетических модификаций является их строгая генетическая детерминированность.
Программированные перестройки хромосом соматических клеток выпадают из поля зрения эволюционистов, так как подобные модификации генома не передаются по наследству. Но если программированные диминуции хроматина и другие хромосомные перестройки связаны с вполне осмысленными действиями ядерного биопроцессора, то нельзя исключить и возможность существования разумной генетической адаптации эукариот, определяющей высокую скорость их эволюцию.
Вместо заключения
embor.embopress.org/content/4/1/18
СПИСОК ЛИТЕРАТУРЫ
1. Патрушев Л.И. Экспрессия генов. — М.,«Наука», 2000 г. – 527 с.
2. Валиев К.А., Кокин Ф.Ф. Из итогов XX века: От кванта к квантовым компьютерам // aakokin.chat.ru/xx.htm
3. Валиев К.А. Квантовая информатика: компьютеры, связь и криптография // Вестник РАН. – 2000 г. – Т. 70, № 8. – с. 688-685.
4. Гаряев П.П., Македонский С.Н., Леонова Е.А. Биокомпьютер на генетических молекулах как реальность // Информационные технологии. – 1997 г. — № 5.
5. Лучинин В.В., Мальцев П.П. Биомолекулы как базис информационных систем будущего // Информационные технологии. – 1997 г. — № 5.
6. Гаряев П.П. Волновой генетический код. — М., 2000 г.
7. Стругацкий А.Н., Стругацкий Б.Н. Попытка к бегству // В сб.: Избранное. – М., “Московский рабочий”, 1989 г. – с. 289-379.
8. Davidson E.H., Britten R.J. Regulation of gene expression: possible role of repetitive sequvences // Sience. – 1979. – Vol.204. – P.1052-1059.
9. Глазков М.В. Петельно-доменная организация генов в эукариотических хромосомах // Молекул. биология. — 1995. — Т.29. – с. 965-982.
10. Miller J.H. Spontaneous mutators in bacteria: insight into pathways of mutagenesis and repair // Annu. Rev. Microbiol. – 1996. – Vol.50. — P.625-643.
11. Льюин Б. Гены / Пер. с англ. — М., “Мир”. – 1987. – 544 с. (с.295-296).
12. Любищев А.А. Проблемы формы систематики эволюции организмов: Сб. статей. — М., «Наука». — 1982. — 278 с.
13. Берг Л.С. Номогенез, или эволюция на основе закономерностей // В сб.: Берг Л.С. Труды по теории эволюции. – 1977. – Л., “Наука”. – С.95-311.
14. Акифьев А.П. Диминуция хроматина – феномен, предназначенный для исследования ряда ключевых вопросов генетики // Вавиловский жернал генетики и селекции. — 1998. — №6 (http://www.bionet.nsc.ru/vogis/vestnik.php?f=1998&p=6_5)
15. Гришанин А.К., Худолий Г.А., Шайхаев Г.О. и др. Генетика. 1996. Т. 32. С. 492.
Комментарии (28)

mahajrod
20.11.2017 21:35+1Гм. Вы бы обновили знания, хотя бы для общего развиятия. Очень много устарешей информации. И список литературы — статья и книги двадцатилетней давности это жесть. Или это стеб такой? Особенно со Стругацкими… притянутых за уши предположений.

genseq Автор
20.11.2017 22:19Спасибо за первую конструктивную критику. Старой (но не устаревшей) информации в ней, действительно, многовато. Например, теория номогенеза была предложена Л.С. Бергом в 1921 году. В развёрнутом виде изложена в его монографии 1922 года. Я же сослался на издание 1977 года. Основные работы А.А. Любищева были написаны в 70-х годах (опубликованы в 1982 году). Жалко, что их сейчас никто не изучает. Ссылку на Стругацких можно причислить к стёбу, но мне очень понравился использованный ими термин «структуральная лингвистика». А воспользоваться этим термином без ссылки на источник было бы некорректно.
Статью написал в 2001 году, но нигде не публиковал. Боялся, что заклюют. Причём как дилетанты (см. выше), так и шизики (см. ниже). Да и сейчас уверен, что заклюют. Надеюсь, что переживу. Или просто перестану сюда наведываться.Zenitchik
20.11.2017 22:29использованный ими термин «структуральная лингвистика»
А определили ли они его?
vconst
21.11.2017 11:10Этот термин определили АБС. Современной науке известен термин «структурная лингвистика» :)
Zenitchik
21.11.2017 12:21Не путайте. Они включили этот термин в текст. Но что на счёт определения?
Про структурную лингвистику я тоже в курсе.genseq Автор
21.11.2017 13:25A Short History of Structural Linguistics
P. Matthews
Professor of Linguistics, University of Cambridge
assets.cambridge.org/97805216/23674/sample/9780521623674ws.pdf
Правильный перевод — структурная лингвистика, но у Стругацких именно «структуральная». И в этом «неправильном» варианте есть своя прелесть. Например, возможность его «незаконного» использования для обозначения структурированности информационных биомолекул.
Кстати, о биологии. Довольно часто даже в научных статьях можно встретить такую неправильную «кальку» с английского языка, как «хромосомальный» (правильно — «хромосомный»).
quqdron
23.11.2017 12:14Напомните откуда?:
Есть на всякий, есть на случай, в корабле, специалист
— наш великий, наш могучий, стуруктуальнийший лингвистgenseq Автор
23.11.2017 12:38www.rusf.ru/abs/konkurs/k_stru00.htm
Аркадий и Борис СТРУГАЦКИЕ
Пусть тахорги в страхе воют,
Издавая визг и писк!
Ведь на них идет войною
Структуральнейший лингвист!
На войне и на дуэли
Получает первый приз –
Символ счастья и веселья –
Структуральнейший лингвист.
Воет ветер дальних странствий,
Раздается жуткий свист –
Это вышел в Подпространство
Структуральнейший лингвист.
Все от ужаса рыдает
И дрожит как банный лист!
Кораблем повелевает
Структуральнейший лингвист.
Есть на всякий, есть на случай
В «Корабле» специалист –
Ваш великий и могучий
Структуральнейший лингвист.
kauri_39
20.11.2017 21:48Статья интересная, но написана языком, сложным для понимания неспециалистами. Как я понял, в ней сделана попытка объяснить "возможность существования разумной генетической адаптации эукариот, определяющей высокую скорость их эволюции". Но загадочную высокую скорость биологической эволюции невозможно объяснить, опираясь на не менее загадочный механизм работы квантовых компьютеров. Для этого надо как минимум объяснить природу мгновенных связей кубитов и посмотреть, не действуют ли они между биомолекулами.
Начинать решение этого вопроса биологам и физикам нужно с ознакомления с общим законом эволюции материи (его вывод — в моей последней публикации). И рассмотреть соответствующий ему вариант мультиверса, который вместе с законом служит основой эволюционной модели мира: http://futurologi.livejournal.com/33983.html
Всё это фантастическое мироздание имеет давно проверенное следствие: мгновенное взаимодействие запутанных частиц (основа работы квантовых компьютеров). Частицы — это системы фотонов, а фотоны — это разумные микровселенные. То есть все частицы внутренне разумны и способны к мгновенному обмену информацией в масштабе своей Вселенной.
Наш разум — это вторичное, постепенно проявляемое свойство разумной материи. И разумная генетическая адаптация эукариот имеет те же корни, что и "разумная адаптация" фундаментальных параметров Вселенной для возможности проведения в ней успешной (полнометражной) эволюции материи. Творцом нашей Вселенной является совокупный Разум всех её кварков, их вселенский "Интернет".
Поэтому уместно уподобить биологическую эволюцию технической. Мысленно посмотрите на Землю в телескоп, в который не видно людей, но видны управляемые ими повозки, корабли, машины, самолёты, растущие города. Вот и в биологической эволюции мы не видим Творца, но видим проявления его творческой инициативы.
Она проявляется в не совсем случайной генерации положительных мутаций в геномах, закрепляемых естественным отбором. Она вроде бы случайна, но протекает в таких неслучайных рамках, что появление в них положительной мутации почти закономерно и дело очень короткого времени. А откатываясь назад к появлению предыдущих неслучайных рамок для успешных эволюций, придём к неслучайным фундаментальным параметрам Вселенной.

liddom
21.11.2017 00:55Трудно убедить себя в том, что наш геном набит вредными для организма эгоистичными нуклеотидными последовательностями, однако это именно так. Пора уходить уже от этого "оранизменного шовинизма". По какой причине вы выделяете организм в единицу, для блага которой должно существовать что-либо? Я понимаю, что это очень привлекательная точка зрения, но тем не менее ошибочная. У организма нет ничего, что качественно выделяли бы его из ряда ген-клетка-организм-популяция-вид-биосфера. Так можно докатился и до размышлений о необходимости нуклеотидных последовательностей в нашем геноме для сверхорганизма Геи.
Почитайте Ричарда Докинза "Эгоистичный ген", а еще лучше "расширенный фенотип" и все встанет на свои места.
Zenitchik
21.11.2017 12:23Не путайте понятия. «Вредный» и «эгоистичный» здесь не синонимы. Мы живы потому что «интересы» большинства наших генов совпадают с нашими (собственно, потому же живы и они).
liddom
21.11.2017 12:29Благодарю за уточнение, согласен с вами. Но, если уж говорить про большинство, то оно скорее "не мешает", чем делает что-то полезное, мне кажется.
Zenitchik
21.11.2017 12:34Да, я неправильно выразился. Надо было написать «большинство работающих генов». Впрочем, бывает, что два гена взаимокомпенсируют вред друг друга — таких чудес в природе тоже хватает.
Megakazbek
21.11.2017 01:47А каким образом может мусорный кусок пропасть из ДНК? Только если он случайно оказался в буквальном смысле смертельно опасен для вида, а иначе он так и продолжит там торчать в будущих поколениях, ведь у него нет никакой пометки для механизма репликации, что он не нужен. Поэтому ничего неожиданного в том, что ДНК заполнена мусором нет, наоборот объяснению подлежали бы случаи, когда мусор в ходе эволюции не возникает или почему-то целенаправленно удаляется.
Zenitchik
21.11.2017 12:23Он может быть случайно вырезан, а поскольку он не важен для организма, такая мутация пройдёт незамеченной.

Mabu
21.11.2017 09:38Хорошо.
Если говорим о разумной и мыслящей ДНК, то для начала нужно ответить на один маленький простой вопрос: что такое сознание?
AkusherA
21.11.2017 09:38Уважаемый genseq, несогласен с заголовком статьи. Нельзя научную теорию называть фантазией;) Очень хорошая статья, но понятна будет до конца только людям с достаточной базой в молекулярной биологии, коих в России не больше сотни… Позволю себе высказать мнение, что ядро скорее представляет жёсткий диск, а процессор находится в цитоплазме, где идёт синтез белков-регуляторов, транскрипционных факторов и т.п., с помощью которых информация из ядра возвращается в виде РНК инструкций. Но, несомненно, ядро — это диск с умным контроллером))) Ещё раз спасибо за статью!!!

fireSparrow
21.11.2017 11:01Основной посыл статьи — «Нет совершенно никаких фактов в поддержку данной теории, но там же в ядре столько всего непонятного и неизвестного происходит, так что — почему бы и нет?»
Какие-то совершенно невразумительные и не понятно откуда высосанные отсылки к квантовому компьютеру, при этом автор, похоже, вообще плохо представляет, чем квантовый компьютер отличается от обычного.
Молекулярные логические операции должны иметь много общего с квантовыми вычислениями и с особенностями работы квантовых компьютеров, к которым можно отнести параллелизм обработки информации, нанометровые размеры логических элементов, а также высокую эффективность вычислений.
С чего бы это они должны? Аргументируйте.
Что вы вообще знаете о квантовых вычислениях? Только то, что у них «параллелизм обработки информации, нанометровые размеры логических элементов, а также высокая эффективность вычислений.»?
Ну и после того, как вы всерьёз делаете отсылки к теории волнового генома, дальше можно было уже вообще не читать.vconst
21.11.2017 13:54Вся статья, какая-то дикая смесь науки и околонаучной ереси, типа дисковых электронов из предыдущей статьи. Особенно доставляет список литературы, где присутствует и упомянутая волновая генетика и даже сами АБС :) Довольно тонкая балансировка между фричеством и прямым троллигном. Очередной забавный курьез, не более того.
genseq Автор
21.11.2017 13:58Насчёт отсылки к теории волнового генома Вы правы. После неё можно и не читать. В своё оправдание могу сказать только то, что около 20 лет назад для меня это было ещё не столь очевидно. Завиральность этой теории особых сомнений не вызывала, но даже в навозной куче встречаются жемчужные зёрна. И в те далёкие времена я всерьёз копался в трудах П.П.Гаряева, пытаясь найти в них хоть что-то ценное. Теперь могу уверенно утверждать, что в его навозной куче жемчуг отсутствует.
С квантовыми вычислениями знаком очень поверхностно. Тут Вы абсолютно правы. Но речь шла не более чем об аналогии с ядерным мышлением. Так что не судите слишком строго. Лучше поищите жемчужные зёрна. Они там есть.
ArtificialLife
Вроде бы, уже давно нет. Где-то читал, что избыточный геном нужен для того, чтобы «важные» гены реже подвергались мутациям.
odu_one
Недавно смотрел лекцию стенфордского университета, где биолог отвечал на этот вопрос. Уже вроде как эта тема закрыта, и механизм действий этого генома и его задачи сейчас хорошо известны
vconst
В том-то и дело, что давно уже не загадка. Назначение «мусорной» ДНК успели выяснить 10 лет спустя, от последней даты используемого автором источника. Не зная очевидных, на данный момент, вещей — автор пытается строить совершенно нелепые предположения на хорошо изученную тему.
genseq Автор
И что там выяснили по поводу назначения «мусорной ДНК»?
vconst
Например: www.youtube.com/watch?v=mBTRC1hyd-g
Не лишним будет и: youtu.be/y7eoXaiLnJs?t=1
genseq Автор
Спасибо за ссылки. Очень «улыбнуло». Правда, вторая у меня не прокачивается, но первую прослушал «от корки до корки». И Вам советую. Детишки в зале, по-моему, были в восторге.
Я слушал Фёдора Кондрашова «живьём» лет 10 назад в соседней лаборатории. Он выступал на семинаре у Симона Эльевича Шноля (своего дедушки). Фёдор ораторствует всё лучше и лучше. Если дослушаете его лекцию до конца, то узнаете, что загадка «мусорной» ДНК так и не была разгадана.
vconst
Что навскидку вспомнилось, то и показал, или вы ожидали двухнедельного исследования и десятки списков литературы? Увы.
Лекции я слушал до конца. Таки про мусорную ДКН, там информации побольше, чем в ваших источниках 20-летней давности. И уж куда больше, чем у АБС или «волновой генетике».
Я вполне допускаю, что в молекулярной биологии вы забыли больше, чем я знал. Но вы зачем-то лезете в квантовые вычисления, в аналоговые компьютеры и прочие темы, в которых совершенно ничего не понимаете, это очень хорошо видно по уровню аналогий, которые вы приводите.
И когда вы пытаетесь сочетать все это вместе, то выходит лютая фоменковщина… Настолько лютая, что непонятно — стеб это или фричество. Вот честное слов — совершенно непонятно.