

Руки для человека являются одними из самых важных конечностей тела. С их помощью мы можем не только орудовать инструментами и получать данные об окружающем мире через тактильные ощущения, но и моделировать движение всего тела (ходьба, бег и т.д.). У многих животных конечностей больше, чему нас, так как у них, как минимум, имеется то, что мы давным-давно утратили, — хвост. Основная функция этой «конечности» заключается в улучшении кинематики тела, т.е. его движения в пространстве. Рыбы плавают в воде, обезьяны лазают по деревьям, птицы маневрируют в воздушных потоках, а ящерицы быстро бегают, ловко ползают и, как оказалось, неплохо «летают». Все это возможно именно благодаря хвосту. Последнее утверждение, конечно, преувеличено, ящерицы не летают, а скорее парят, перепрыгивая с места на место. И как установили ученые из Калифорнийского университета в Беркли (США), хвост играет важную роль в успешности приземлений на вертикальные поверхности. Данный вывод является итогом наблюдений, математического моделирования и даже тестов с применением геккона-робота. Какими же механическими свойствами обладает хвост геккона, как именно он помогает избежать крушения при посадке, и какой бы была посадка без хвоста? Ответы на эти вопросы мы найдем в докладе ученых. Полетели. Поехали.
Основа исследования
Рептилии ассоциируются у человека с существами, которые являются живым примером фразы «рожденный ползать летать не сможет». От того они и получили свое второе название — пресмыкающиеся. Однако, как показывает практика (Чешуйчатый Икар: кинематика «полета» древесной змеи), ползать это далеко не единственный манёвр, на который способны рептилии.

Шлемоносный василиск, которого прозвали ящерицей Иисуса Христа за удивительную способность бегать по воде.
Конечно же, помимо нестандартных полетов и бега на короткие дистанции по воде, рептилии умеют отлично лазать по самым разным поверхностям. Одними из самых умелых в этом навыке считаются гекконы. Возможно это благодаря необычному строению их лапок, покрытых множеством мелких волосков, увеличивающих сцепление с поверхностью.

РЭМ снимок ступни геккона.
При этом хвост также задействован. Наблюдения за азиатскими плоскохвостыми гекконами (Hemidactylus platyurus) показали, что во время вертикального бега и контакте лап со скользкой поверхностью гекконы резко упираются хвостом в поверхность, предотвращая падение. На этот манёвр им требуется не более 30 миллисекунд. Если же геккону не удалось удержаться, и он все таки упал, то во время падения его тело принимает позу парашютиста (конечности во все стороны). При падении спиной к земле гекконы за 100 мс способны перевернуться, совершив для этого взмах хвостом. Не говоря уже о том, что именно хвост позволяет некоторым видам гекконов бегать по воде, о чем мы уже говорили ранее (Run, Gecko, run: гибридный механизм движения по воде у гекконов).

Азиатский плоскохвостый геккон (Hemidactylus platyurus).
Этих примеров уже достаточно, чтобы сделать очевидный вывод — хвост для гекконов (и для рептилий в целом) является крайне важной частью тела. Но ученые не были бы учеными, если бы не решили копать дальше.
Существует теория, что хвост необходим для стабилизации и управления во время планирования. Чтобы это проверить, ученые засунули гекконов в аэродинамическую трубу. Важно отметить, что в этих опытах брали участие гекконы, которым не присуще планирование в условиях дикой природы. Несмотря на это, рептилии успешно выполняли контролируемое равновесное скольжение (т.е. с линейной траекторией относительно потока воздуха). При этом было установлено, что круговые движения хвоста сопровождались поворотом тела. Планирующие гекконы, которые вращали свой частично изогнутый хвост по часовой или против часовой стрелки инициировали повороты вправо или влево (т.е. рыскание). Учитывая закон сохранения углового момента, вращение хвоста в одном направлении приводит к повороту тела в другом, тем самым вызывая рыскание.

Рыскание на примере модели самолета.
Учитывая эти данные, ученые предположили, что азиатский плоскохвостый геккон (Hemidactylus platyurus) не только способен планировать в воздухе, но и может использовать свой хвост для поворота и маневрирования к месту посадки.
Но одной только гипотезы недостаточно, ее необходимо было подтвердить. Для этого были проведены наблюдения за живыми гекконами, математическое моделирование и опыты с применением гекконов-роботов.
Результаты исследования
Авторы исследования отмечают, что среди моделей поведения в воздухе, исследованных на сегодняшний день, конечная стадия направленного воздушного снижения или планирования является наименее изученной. Особенно в аспекте поглощения энергии при ударе у планеров с ограниченными возможностями аэродинамического управления. У птиц (насекомых, летучих мышей и т.д.) сила посадки может быть значительно ниже силы взлета. Уменьшение кинетической энергии перед приземлением или посадкой в основном достигается за счет взмахов крыльев.
В отличие от крылатых летунов, животые-планеры (белки-летяги, змеи, лягушки, ящерицы, кагуаны и т.д.) движутся к точке приземления с достаточно большой скоростью и могут испытывать большие пиковые силы при приземлении, используя свои конечности и тело.
Летучий дракон не убегает от хищников, он от них улетает.
К примеру, летающие ящерицы из рода летучих драконов (Draco) вытягивают свои ребра для достижения высокой скорости планирования и могут ориентировать свое тело почти параллельно поверхности непосредственно перед приземлением всеми конечностями одновременно, чтобы замедлиться и облегчить контакт с поверхностью.
Пилохвостая ящерица (Holaspis guentheri).
А вот пилохвостая ящерица (Holaspis guentheri), несмотря на свою способность сплющивать дорсовентральную часть тела, вместо этого использует преимущества низких нагрузок на крыло* за счет уменьшения плотности скелета для создания нужной траектории спуска.
Удельная нагрузка на крыло* — отношение массы летящего тела к площади несущей поверхности.Но вот у азиатских плоскохвостых гекконов нагрузка на крыло очень высока (~ 65 Н/м2) по сравнению с большинством планеров, даже с теми, у кого масса тела значительно больше их.
Анализ записей планирования гекконов показал, что их угол скольжения был почти в два раза больше, а коэффициент планирования почти вдвое меньше, чем у естественных планеров (имеющих специальные физиологические особенности, нужные для планирования), таких как летучие драконы.
В результате азиатские плоскохвостые гекконы продемонстрировали баллистическое неустойчивое планирование на коротких дистанциях, крайне быстрое приземление и весьма сильный удар (видео ниже).
Видео №1: планирование и приземление гекконов в естественной среде обитания.
Гекконы ускорялись на протяжение всего планирования и достигали равновесия лишь в конце полета (4 из 21 особей ускорялись вплоть до удара о поверхность; 1b).

Изображение №1
Тем не менее более половины испытуемых (13 из 21) достигали точки приземления (1a). Учитывая, что H. platyurus не является естественным планером (грубо говоря), его полеты были баллистическим (т.е. без подъемной силы) и короткими. При этом скорость приземления головой вперед была 6.0 ± 0.9 м/с. Во время полета можно было заметить изменения положения тела геккона, которые учащались по мере приближения к точке посадки. При этом угол приближения оставался практически неизменным — 53 ± 5.8 ° (1d), как и у естественных планеров (52° у Ptychozoon kuhli и 57° у P. lionatum).
График скорости в зависимости от продолжительности планирования (1b) показывает, что по мере приближения к дереву (место посадки) гекконы снижали скорость, т.е. пытались выполнить некий специальный манёвр приземления. Однако у гекконов отсутствует специальная аэродинамическая морфология (выраженные перепонки на лапках, кожные складки вдоль тела и т.д.), от чего 4 из 21 особей продолжали ускорение вплоть до столкновения, а остальные смогли замедлиться лишь на 6.4 ± 4.9% от максимальной скорости полета. Следовательно, они должны были поглотить почти весь свой полетный импульс при ударе, а это должно приводить к травмам или падениям.
Любопытным является то, что некоторые гекконы обладают необычным строением черепа, снижающим вероятность получения травмы ввиду не очень умелого планирования.
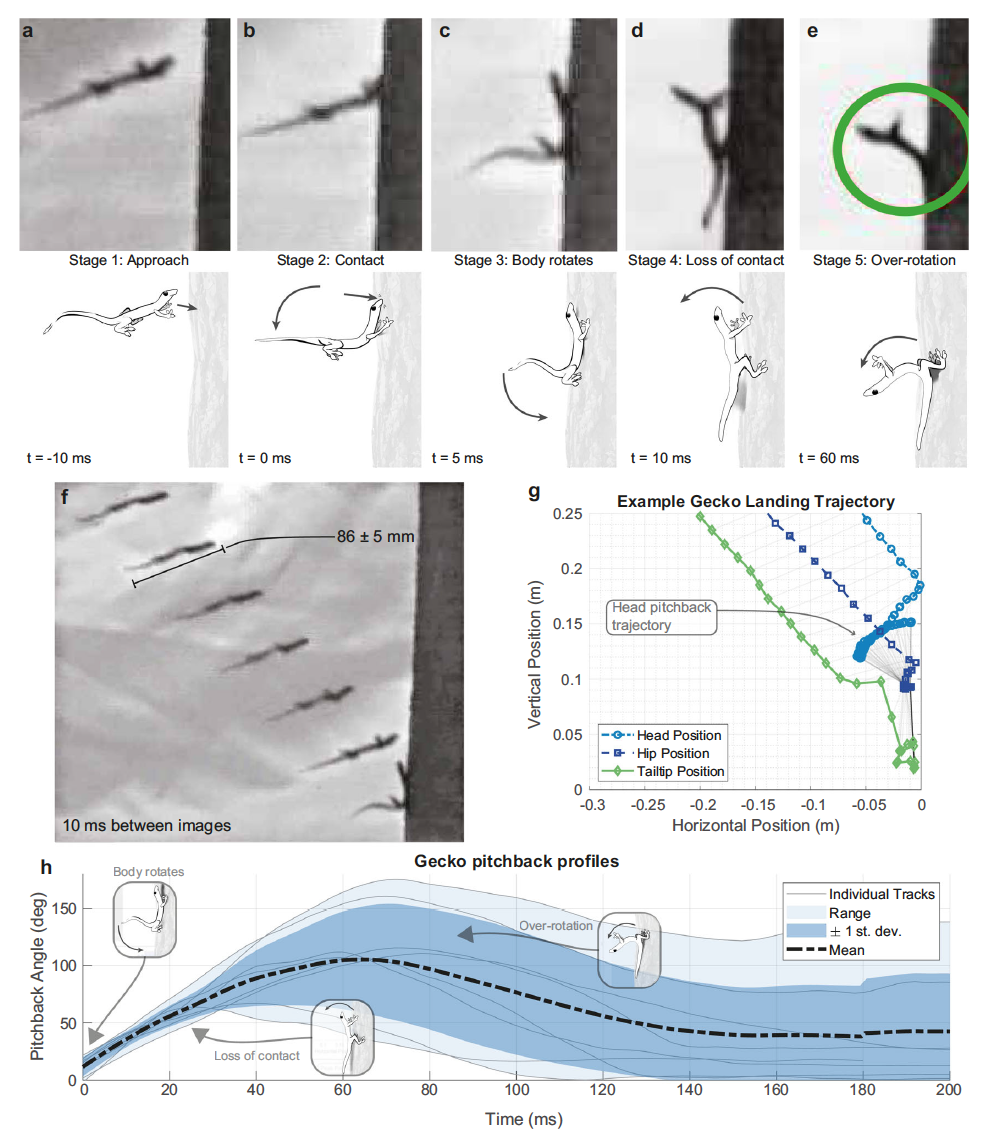
Изображение №2
Наблюдения в условиях естественной среды обитания показали, что гекконы используют хвост и задние лапки, чтобы безопасно приземлиться на ствол дерева (2a-2f).
Траектория головой вперед сопряжена с большим угловым моментом (моментом импульса) по тангажу. Используя хвост для создания длинного плеча импульса, геккон может постепенно рассеивать этот импульс, отклоняясь назад, и в конечном итоге успешно приземлиться с уменьшенной силой. Ученые решили дать этому манёвру название «реакция задержки падения» (FAR от fall arrest response).

Тангаж на примере модели самолета.
В дополнение к 21 планированию, записанному на камеру с большого расстояния, были сделаны записи с малого растения еще 16 планирований (1c и 1d). Из них пять особей успешно контактировали с поверхностью точки приземления всеми лапами. Одна особь упала с дерева, качнувшись назад, тем самым потеряв сцепление задних лап с поверхностью.
Когда особь (8 из 16 особей) теряла сцепление с поверхностью передними лапами, наблюдался FAR манёвр с отклонением тела назад и упором на хвост, что позволяло им успешно приземлиться и удержаться на дереве. Бесхвостая (по неизвестным причинам) особь, теряя контакт передними лапами с поверхностью, не могла использовать хвост для стабилизации тела, в результате чего падала (2d и 2e). Это лишний раз подтверждает важность хвоста в успешности приземления после планирования.
Манёвр приземления геккона ученые разделили на пять этапов. Во-первых, планирующие гекконы достигли цели, наклонив тело вверх на 16 ± 8.4° по горизонтали (2g). Когда геккон завершал спуск по воздуху (этап 1; 2а), начальный контакт происходил с его головой и передней частью туловища, поглощающей кинетическую энергию (этап 2; 2b).
Столкновение увеличивало угловой момент, который вызвал вращение туловища вниз по направлению к стволу дерева. Затем задние лапы контактировали с вертикальной поверхностью (этап 3; 2c). Далее обе передние лапы начинали превышать предел сцепления и терять контакт с поверхностью (этап 4; 2d).
На заключительном этапе отклонение назад с прижатым к дереву хвостом предотвратить было уже невозможно, и туловище геккона начинало поворачиваться (отклоняться) назад от дерева (этап 5; 2e).
Во время наблюдений за гекконами были получены углы отклонения тела (2h), составляющие до 175° (в среднем 114 ± 16°). Изображение 2e отлично показывает положение тела в момент приземления и его сопутствующего отклонения от дерева.
Ученые отмечают, что во время бега по стенам время между проскальзыванием передней лапы и касанием хвоста к поверхности составляло 47 мс. В FAR время, затрачиваемое между потерей контакта передней лапы и пиковым наклоном назад, составляло примерно 64 мс (2h). То есть времени вполне достаточно, чтобы геккон мог задействовать свой хвост, несмотря на высокую скорость планирования и приземления.
На 2h показано угловое положение тела (θ) как функция времени на всем протяжении FAR манёвра. Гекконы демонстрировали значительный наклон туловища от дерева со скоростью от 2057 ± 762 °/с до среднего угла 114 ± 16 ° по отношению к лесной подстилке (т.е. к горизонтали). Средняя продолжительность всего манёвра с учетом наклона тела и восстановлением контакта с поверхностью составляет 138 ± 15 мс.
Дабы лучше понять влияние хвоста на степень успешности приземления, учеными была создана упрощенная модель динамики твердого тела (т.е. тела геккона) во время тангажа в ходе FAR маневрирования.

Изображение №3
Из видео, записанного в тропическом лесу, видно, что задние лапы геккона не регрессируют и не соскальзывают во время манёвра с наклоном назад. Для количественной оценки FAR задние лапы были представлены в виде шарнирного соединения, которое демонстрирует компонент силы реакции, перпендикулярной и касательной к вертикальной поверхности дерева (изображение №3). Туловище геккона в модели было представлено одним твердым однородным объектом длиной LB, вращающимся вокруг этого шарнира (синий овал на изображении №3).
Если сравнивать ступни гекконов и их хвост, но у хвоста сцепление с поверхностью на несколько порядков ниже, так как на вентральной поверхности нет специальных волосков, как на ступнях. Кроме того, трение хвоста может дать туловищу лишь минимальный крутящий момент из-за небольшого плеча момента между ним и точкой поворота на задних лапах.
Следовательно, ученые предложили упростить модель, предполагая, что сила трения между хвостом и деревом будет минимальным. Посему сила хвоста (FT) представлена в виде точечной нагрузки, перпендикулярной вертикальной поверхности дерева, расположенной на расстоянии LT от шарнирного соединения, через которое силы могут передаваться на туловище.
Учитывая предложенный вариант модели, сила задних лап (FF), необходимая для удержания ящерицы на вертикальной поверхности, будет обратно пропорциональна длине хвоста. Это уже указывает на важность хвоста, но не дает полного понимания того, что состояние максимального отклонения тела от вертикальной поверхности является критической точкой, после которой задние лапы могут потерять контакт с деревом.
Чтобы исследовать силы лап на протяжении всего манёвра с наклоном назад, ученые предложили исследовать три отдельных случая, для которых возможны простые численные решения уравнений движения: постоянное угловое замедление (θ = константа); постоянная сила хвоста (FT = постоянная); пропорциональная сила хвоста (FT/θ = константа).
В условиях модели физически постоянное замедление будет представлять собой минимальный крутящий момент в точке поворота и наиболее постепенное замедление при движении по тангажу. Постоянная сила хвоста представляет собой максимальную механическую работу, которая может быть применена, если сила хвоста ограничивается его мускулатурой. А пропорциональная сила хвоста будет представлять собой упругую реакцию тканей тела.
Для каждого из этих случаев выводилась сила тангажа (наклона назад), используя начальную угловую скорость, измеренную во время среднего тангажа (2h) в качестве начального условия. Результаты данного анализа представлены на изображениях 4a-4c.

Изображение №4
Из анализа становится очевидно, что самая большая составляющая силы в каждом случае связана с угловым замедлением тела, которое представляет собой геккона, рассеивающего кинетическую энергию, накопленную во время его планирования. Сравнение прогнозируемого тангажа с наблюдениями (4d и 4e) показало соответствие как пикового угла тангажа, так и продолжительности в случае постоянного замедления.
Продолжив анализ, ученые смогли также спрогнозировать среднюю силу хвоста по диапазону значений его длины (4f). Была обнаружена зависимость от обратной пропорциональности. Анализ с учетом хвоста после автотомии (самостоятельное отбрасывание хвоста) показал, что такая малая длина приводит к значительному и резкому увеличению силы, необходимой задним лапам для удержания на поверхности. Другими словами, эти данные подтверждают факт того, что бесхвостые особи значительно хуже справляются с приземлением после планирования.
Согласно этому выводу, для бесхвостых гекконов вида H. platyurus сила лап, необходимая для удержания, должна быть в 5 раз выше, чем сила у гекконов с целым хвостом (4f). Это дополнительно подтверждается полевыми наблюдениями, когда около 87% гекконов с хвостами успешно приземлялись на вертикальную поверхность дерева, тогда как бесхвостые практически всегда падали.
Результатов математического моделирования вполне достаточно, чтобы служить подтверждением теории, что хвост гекконам помогает стабилизировать приземление. Однако ученые решили пойти еще дальше и создали роботизированную модель геккона для дальнейшего анализа.
В ходе опытов с роботом измерялись силы, возникающие при «запуске» робота на вертикальную поверхность с помощью катапульты при заданных скоростях от 3 до 5 м/с и углах приближения, полученных из полевых наблюдений (1c и 1d).

Изображение №5
Было проведено 79 экспериментов (5a-5d). Результаты показали количественную и качественную схожесть поведения робота, математической модели и живых гекконов (5e-5i).
Видео №2: приземление роботизированного геккона.
Анализ механики контакта при приземлении позволил понять, почему хвостовой манёвр так важен. Когда передние лапы касаются стены, поступательный момент модели робота быстро преобразуется в угловой момент. Следовательно, когда задние лапы модели робота достигают поверхности, модель будет продолжать вращаться, а передние лапы отсоединятся, если сила сцепления недостаточна. Затем модель робота поворачивается от вертикальной поверхности к земле настолько, насколько позволяет ее геометрия. На этом этапе робот без хвоста с большой долей вероятности теряет сцепление задних лап с поверхностью. Но вот наличие хвоста обеспечивает сопротивление крену тела назад и действует как амортизатор. На графиках 5j представлена кинематика, наблюдаемая на видеозаписях робота.
На 5k показаны совокупные результаты 26 запусков с успешным восстановлением после тангажа. Модель робота показала среднюю начальную скорость наклона по тангажу 1438 °/с по сравнению с 2057 °/с у живых гекконов.
Были обнаружены аналогичные пиковые углы наклона, но более медленная скорость восстановления. Роботу для восстановления требовалось примерно в три раза больше времени, чем для достижения максимального угла, в отличие от более симметричного FAR, наблюдаемого у гекконов. Это объясняется используемыми в разработке робота материалами, лишенного мускульной морфологии реального геккона (4e).
Во время следующих опытов с роботом применялись датчики силы, записывающие силы лап во время приземления. В ходе этих экспериментов робота запускали под углом 45°. После чего анализировалась роль и эффективность хвостового манёвра, инициируемого контактным переключателем.
Видео №3: примеры приземления роботизированного геккона с хвостовым манёвром и без хвоста вообще.

Изображение №6
Результаты опытов (6a-6c) показывают, что укороченный хвост значительно увеличил нагрузку на лапы, что согласуется с предсказаниями математической модели и наблюдаемыми за живыми гекконами. По всем трем длинам хвоста наблюдались согласованные профили наклона спины, указывающие на то, что увеличение силы в основном связано с изменением длины хвоста, а не с влиянием потери хвоста на динамику туловища. На 6d и 6h виден эффект хвостового манёвра при наличии полноценного хвоста: контакт передних лап с поверхностью инициировал закручивание хвоста вниз, управляемое сервоприводом. Было установлено, что в тестах с хвостовым манёвром снижение силы лап было более резким, чем в случаях с пассивным хвостом (без привода).
Как и ожидалось, в опытах по всем диапазонам углов и скоростей приближения роботы с хвостом превзошли бесхвостых, что полностью согласуется с наблюдениями за гекконами. Модель бесхвостого робота смогла успешно приземлиться только в 15% испытаний (n = 20) по сравнению с 55% при наличии хвоста (n = 38) (7a).

Изображение №7
При вычислении значения силы лап роботов из предыдущего опыта было обнаружено, что хвост уменьшает силу во время замедления (7b). Это согласуется с математической моделью, т.е. сила лап увеличивается обратно пропорционально длине хоста.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В данном труде ученые решили рассмотреть «полет» налетающего существа, а именно азиатского плоскохвостого геккона. Эти маленькие ящерицы, хоть и не имеют никаких физиологических приспособлений, вполне неплохо планируют с дерева на дерево, в чем им сильно помогает хвост.
Проведенные наблюдения, математическое моделирование и опыты с роботизированными гекконами показали, что хвост позволяет не только стабилизировать сам процесс планирования, но и приземление на вертикальную поверхность. Учитывая отсутствие крыльев или хотя бы кожных складок вдоль тела, планирование гекконов протекает на весьма больших скоростях и заканчивается жесткой посадкой, когда они буквально врезаются в дерево головой вперед. Сила удара такова, что их тело начинает отклоняться назад, а передние лапы теряют сцепление с поверхностью. При наличии хвоста она способны нивелировать этот процесс и стабилизировать положение тела. Если же хвоста нет, то сцепление теряют и задние лапы, а геккон отправляется в свободное падение на лесную подстилку. При этом одного лишь наличия хвоста может оказаться недостаточно, потому геккон совершает им определенные движения, позволяющие реализовать успешную посадку.
Ученые отмечают, что подобное поведение является ярким примером того, что эволюция не всегда идет по логическому пути. Порой приходится работать с тем, что имеется. Одни виды, ввиду более активного использования воздушного пространства, предпочли наличие более выраженных перепонок между пальпации лап или специальных крылообразных складок вдоль тела. Но исследуемые в данном труде гекконы не особо нуждаются в регулярных рейсах, а потому остановились на крепком черепе и очень подвижном хвосте.
Гекконы показывают, что для решения какой-либо задачи не нужны идеальные инструменты и условия, иногда достаточно того, что есть, плюс немного креативности и смекалки (а еще крепкий череп, про это тоже не стоит забывать).
Пятничный офф-топ:
Иногда, чтобы победить хищника, его нужно просто сильно озадачить.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?