

Продолжительность жизни дрозофилы составляет примерно 3 месяца. Официальный старожил-рекордсмен (чей возраст был подтвержден документально) Жанна Кальман прожила 122 года (1875-1997). Возраст губки вида Anoxycalyx joubini, вычисленный Томасом Брейем по данным потребления кислорода, составляет порядка 10000 лет. Но все эти цифры меркнут на фоне возраста самой планеты Земля, которая сформировалась примерно 4.5 миллиардов лет тому назад. Первая же жизнь зародилась немногим позже, примерно 4.1 млрд. лет назад. Ученый свет уже очень давно ведет ожесточенные споры на тему того, как, где и при каких обстоятельствах произошло это воистину важнейшее событие в истории планеты. Какими бы ни были теории о возникновении жизни, понятно одно — первые клетки должны были научиться создавать компартменты (отсеки), дабы облегчить первые химические реакции. Но как это произошло? Ученые из Мюнхенского университета (Германия) предполагают, что рост и деление безмембранных микрокапель мог возникнуть в газосодержащих порах нагретой породы, которой на ранней Земле было предостаточно. В чем особенность этих пор, как именно там могли зародиться первые полноценные клетки, и что этому способствовало? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Клетки самых разных организмов, от сложных (к примеру, человек) до самых простых (к примеру, инфузория-туфелька), могут отличаться по своему составу, но быть схожими по структуре. Клетка содержит ряд органелл (митохондрии, хлоропласты, пероксисомы, лизосомы, эндоплазматический ретикулум, ядро и аппарат Гольджи), которых можно назвать компартментами. Компартментализация — это разделение клеток эукариот на отсеки (компартменты), покрытые оболочкой, где протекают различные биохимические процессы.
Считается, что именно компартментализация является важнейшей составляющей зарождения жизни за счет пространственной локализации молекул и облегчения первых химических реакций. Один из возможных путей компартментализации является разделение жидкой фазы противоположно заряженных полиэлектролитов в водном растворе. Этот процесс приводит к образованию безмембранных химически обогащенных капель.
Эти микрокапли коацервата (первичного бульона) представляют собой модели протоклеток, поскольку они формируются с незначительной химической идентичностью в широком диапазоне физико-химических условий. Они локализуют и концентрируют ряд различных молекул и проявляют молекулярную селективность путем разделения. Кроме того, капли коацервата способствуют сборке бислоев жирных кислот на их внешней поверхности и легко поддерживают каталитические реакции, такие как примитивный РНК-катализ. Это обеспечивает путь к мембраносвязанной компартментализации, наблюдаемой в современных клетках.
Ученые отмечают, что события слияния, деление и поддержание протоклеток коацервата были бы крайне важны для эволюции компартментализированных молекул.
Слияние и рост протоклеток необходимы для обмена молекулами и генетическим материалом. В растворе капли коацервата будут иметь тенденцию к коалесценции (слияние частиц), в конечном итоге образуя объемную макрофазу коацервата, что ограничивает их роль в качестве протоклеток.
Разделение протоклеток коацерватов необходимо для передачи молекулярной информации последующим дочерним протоклеткам, которые могут передать эволюционные преимущества следующему поколению.
Для достижения деления современные клетки используют сложный механизм, в котором задействованы регуляторные белки, каркасные белки, ферменты и химические посредники. Но в пребиотическом мире разделение должно было основываться на других факторах. Некоторые исследования предполагают, что деление липидных везикул могло быть вызвано осмотическими изменениями, химическими изменениями, температурой и сдвигающими силами.
При этом о делении химически обогащенных безмембранных коацерватных протоклеток известно очень мало. Есть теория, что почкование химически активных безмембранных капель достигается за счет потока субстрата и продукта через границу раздела, которая находится в определенном режиме поверхностного натяжения. Но это лишь теория, которую пока никто на практике не подтвердил.
И тут ученые обратили внимание на поры. Известно, что капиллярные протоки, вызванные тепловыми потоками в порах миллиметрового размера, аккумулируют молекулы в зависимости от их размера на границе раздела газ-вода газовых включений. Моделирование и эксперименты показывают, что на границу раздела действуют две основные силы: капиллярные потоки от холодной стороны к теплой и пертурбативные потоки. Эти силы вызывают быстрое движение частиц, приводя к их контакту и слиянию. В таких условиях липидные молекулы накапливаются на границе раздела, создавая везикулярные структуры и подвергаясь делению под действием потоков Марангони* и конвекции.
Эффект Марангони* — явление переноса вещества вдоль границы раздела (интерфейса) двух сред, возникающее вследствие наличия градиента поверхностного натяжения.Следовательно, эти исследования показывают, что ростом, делением и поддержанием капель коацервата можно управлять с помощью физических потоков внутри термальных порах.
В рассматриваемом нами сегодня труде ученые изучают влияние неравновесных условий, создаваемых нагретыми порами, содержащими пузырьки газа, на механизмы роста и деления сложных микрокапель коацервата, образованных при смешивании полианионных соединений (карбоксиметилдекстран (CM-Dex), аденозин-5’-трифосфат (ATD)) и поликатионных соединений (полидиаллилдиметиламмонийхлорид (PDDA), поли-1-лизин (pLys)).
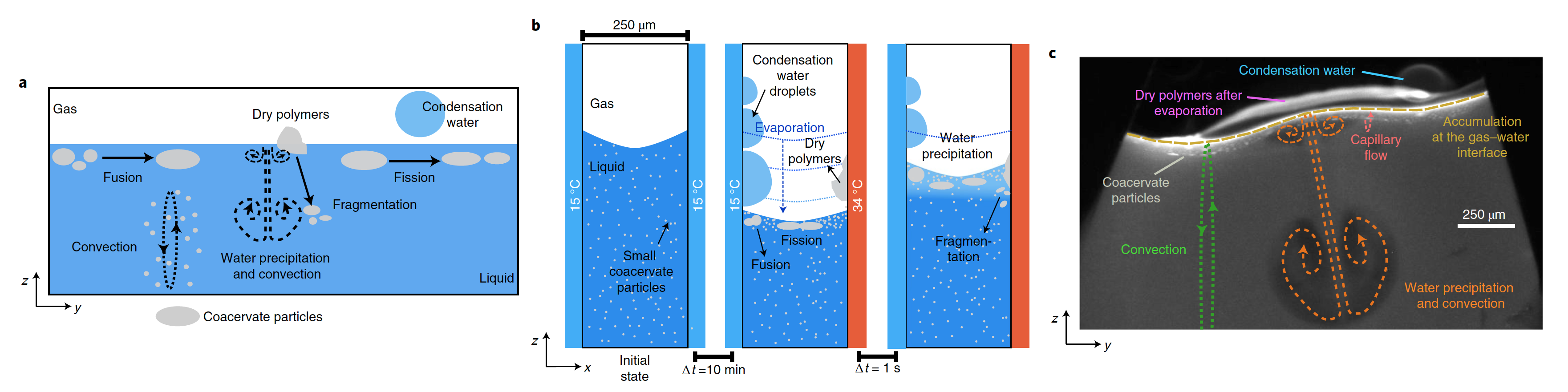
Исследование показало, что накопление компонентов, образующих коацерват, на границе раздела газ-вода газового пузыря приводит к росту за счет слияния микрокапель коацервата. Важное наблюдение заключается в том, что микрожидкостный водный круговорот, индуцированный тепловым градиентом, создает пертурбативные потоки на границе раздела газ-вода, которые приводят к делению и фрагментации капель коацервата с использованием чисто физических процессов (1a-1c).
Изображение №1
Это является прямым доказательством того, что физических сил в замкнутой среде достаточно для обеспечения механизма безмембранного деления протоклеток без сложных механизмов или химических реакций.
Результаты исследования
Чтобы охарактеризовать влияние неравновесных возмущений на микрокапли коацервата, ученые экспериментально воссоздали поры нагретой породы, заполненные пузырьками жидкости и газа. Лист политетрафторэтилена (ПТФЭ) толщиной 250 мкм с острыми треугольными структурами помещался между оптически прозрачным сапфиром и пластиной из диоксида кремния (2а).

Изображение №2
Жидкость загружалась в полость через микрожидкостные трубки, а пузырьки газа создавались путем неполного заполнения жидкостью треугольных полостей (2b). Образцы помещались под специально изготовленный микроскоп, где градиент температуры создавался за счет дифференцированного нагрева сапфира стержневыми резисторами, вставленными в медный держатель, и за счет охлаждения медной задней пластины через соединение с водяной баней (2c и 2d). Градиенты температуры варьировались от 15 до 29 °C с точностью ± 1 °C. Визуализация велась через прозрачный сапфир, а камера фокусировалась на холодной стене.
Дисперсные системы* микрокапель коацервата были получены путем смешивания отрицательно заряженных модифицированных сахаров CM-Dex или ATP с положительно заряженными полиэлектролитами, т.е. pLys или PDDA (2e). Смеси CM-Dex:PDDA и CM-Dex:pLys получали при молярных соотношениях 6:1 и 4:1. А смеси ATP:PDDA и ATP:pLys получали при молярном соотношении 4:1. Общие концентрации полимера варьировали от 2 до 20 мМ.
Дисперсная система* — образования из двух или более фаз (тел), которые практически не смешиваются и не реагируют друг с другом химически.Начальная концентрация определяла плотность капель коацервата в дисперсии и конечное количество материала, накапливающегося на границе раздела газ-вода. Чтобы визуализировать капли коацервата, использовался 0.1% флуоресцеинизотиоцианат (FITC), меченный CM-Dex или pLys. Дисперсии коацервата готовились либо в растворе 0.1 М натрия + бициновый буфер, либо в растворе 10 мМ C4H11NO3 + 4 мМ MgCl2. Контрольные эксперименты показали, что не было заметной разницы между двумя разными буферами в отношении динамики коацервата в тепловой ловушке.
При загрузке дисперсной системы коацервата (20 мМ CM-Dex:PDDA в 0.1 М Na + бициновый буфер) в тепловую ловушку визуализация (снимки делались каждую ~ 1 с) показала равномерное присутствие мелких капель коацервата (< 10 мкм), рассредоточенных по полости (3а).

Изображение №3
После дифференциального нагрева с двух сторон ловушки (теплая сторона 49 °C; холодная сторона 20 °C) флуоресцентные капли испытали конвективные потоки в объеме раствора. Скорость конвективного потока можно было регулировать за счет изменения разницы температур.
В ходе наблюдений было замечено, что капли коацервата в объеме раствора переносились конвекционным потоком к границе раздела газ-вода, где они накапливались и начинали расти за счет слияния (3b и 3c; видео №1). На границе раздела капли двигались параллельно ей, за счет чего возникал контакт между ними с последующим слиянием.
Видео №1: слияние капель.
Процесс слияния двух капель коацервата занимал несколько секунд (от 1 до 10 с) и приводил к образованию коацерватов эллиптической формы (3d — слияние трех капель).
Рост коацерватов с течением времени количественно оценивался по изображениям оптической микроскопии. Анализ показал, что коацерваты CM-Dex:PDDA достигали максимального размера в 150 мкм. Эксперименты с другим буфером (10 мМ C4H11NO3 + 4 мМ MgCl2) или разными полимерами с разной молекулярной массой (CM-Dex:pLys, ATP:pLys, ATP:PDDA или CM-Dex:pLys с более высокой молекулярной массой) показали аналогичное поведение с небольшими различиями в конечном размере коацервата (3e).
Дополнительно была проведена оценка влияния общей концентрации полимера на скорость роста и конечный размер капель коацервата. Для этого были проведены эксперименты с постоянным температурным градиентом (теплая сторона 49 °C и холодная сторона 20 °C), постоянным буфером (10 мМ C4H11NO3 + 4 мМ MgCl2) и полимерами (молярное соотношение CM-Dex:PDDA было 6:1).
Полученных в ходе визуализации изображения анализировались с помощью ImageJ или LabVIEW для определения скорости роста и среднего размера капель спустя час воздействия температурного градиента на границу раздела.
Анализ данных показал, что на конечный размер капель и скорость роста исходная концентрация полимера существенно не влияла. Также было замечено, что различия в размере капель между экспериментами были весьма существенны. По мнению ученых, это может быть связано с флуктуациями солей, вызванными микромасштабным круговоротом воды, а также с внутренней стохастической природой слияния капель.
Несмотря на это, во всех случаях (более 50 экспериментов с разными условиями) капли коацервата накапливались и сливались вместе. Это указывает на то, что накопление, слияние и поддержание капли коацервата в газе на границе раздела газ-вода — это общее явление, обусловленное силами в тепловой ловушке, а не химическим составом дисперсной системы коацервата.
Дополнительно были проведены опыты с исходными концентрациями полимера ниже критической концентрации коацервата (CCC от critical coacervate concentration), т.е. менее 1 мМ в дисперсной системе коацервата CM-Dex: PDDA.
При исходной концентрации 0.2 или 0.05 мМ капли коацервата обнаружить с помощью микроскопии не удавалось, несмотря на очевидное повышение концентрации полимера на границе раздела газ-вода. Эти результаты показывают, что термические поры действуют на механическом уровне, управляя слиянием ранее существовавших капель коацервата, за которым следует деление капель путем растяжения/фрагментации и агрегации.
Необходимо было удостовериться, что граница раздела газ-вода являются важной составляющей выше описанных процессов. Для этого были проведены опыты, в которых полости не содержали газа.
Изображения, полученные с помощью оптической микроскопии с временным разрешением, показали, что объемные капли коацервата (< 15 мкм) переносились в полости конвекционным потоком со скоростью около 1.6 ± 0.4 мкм/с, но не подвергались процессам слияния.
Если тепловой поток отсутствовал, то практически все капли оседали на дно полости, а затем сливались в одну большую каплю. При наличии температурного градиента конвекционный поток в объеме предотвращал осаждение капель коацервата, поддерживая их в термофоретическом потоке. Чем больше был градиент, тем меньше капель подвергалось осаждению в камере без газа. Если же газ был, а значит и граница раздела газ-вода, то для достижения такого же эффекта достаточно было и минимального градиента температур.
Вышеописанные результаты подтверждают, что потоки на границе раздела газ-вода приводят к накоплению капель коацервата на границе раздела, событиям слияния между каплями и к удержанию капель от осаждения. В отсутствие теплового потока капли оседают на дно полости. Следовательно, комбинация конвекции и капиллярного потока на границе раздела удерживает капли на границе раздела газ-вода или сопутствует циркуляции внутри полости в течение длительных периодов времени.
Данные наблюдений также показали, что противодействующие силы на границе раздела приводят к удлинению капель (3c и 3d). Поскольку эллиптическая форма связана с начальными стадиями деления везикул, возник вопрос — могут ли силы, возникающие внутри тестовой полости, быть достаточно существенными, чтобы привести к делению безмембранной капли коацервата.
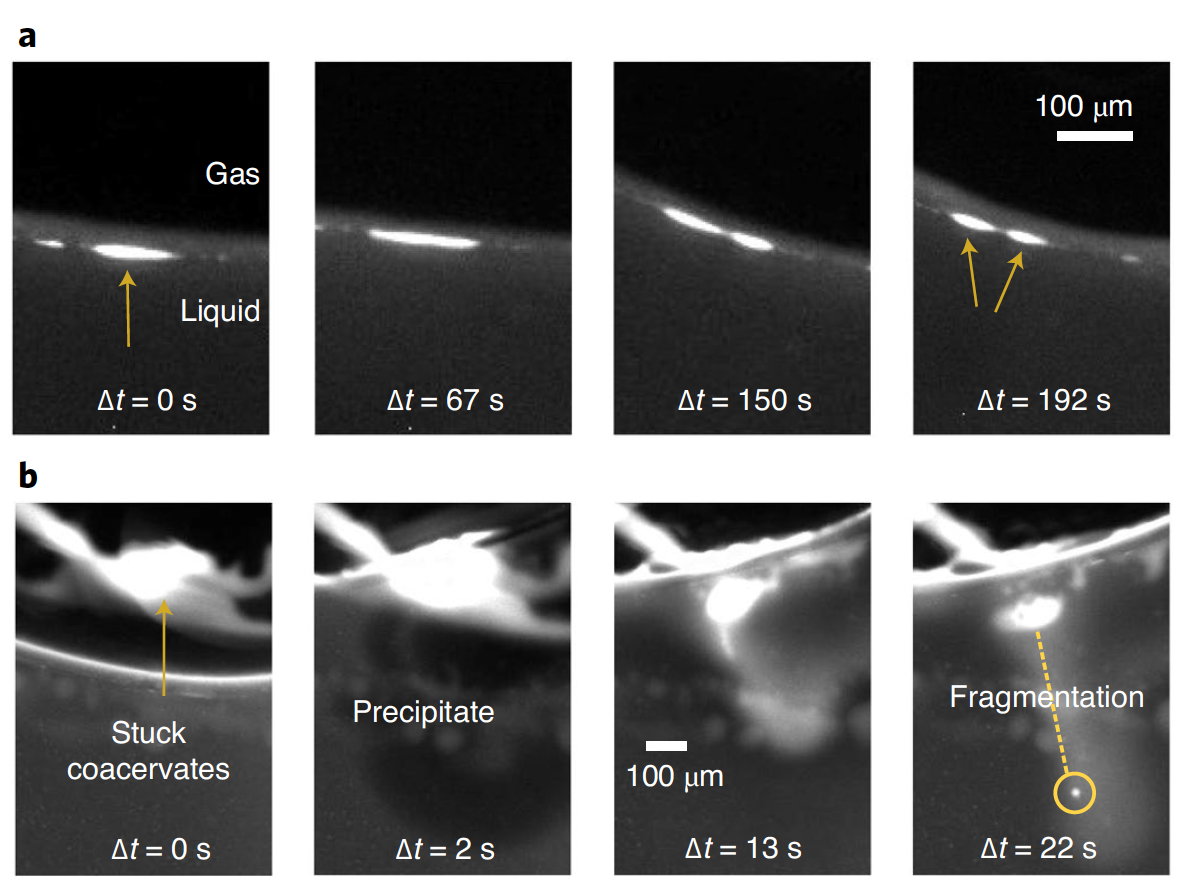
К дисперсной системе CM-Dex:PDDA был применен температурный градиент 34 °C / 15 °C (теплая и холодная стенки камеры). Микроскопия показала, что капли коацервата накапливались, сливались и становились эллиптически вытянутыми на границе раздела газ-вода (3c и 3d). При накоплении капли постепенно вытягивались вдоль границы раздела, пока не разделялись с образованием двух дочерних протоклеток аналогичного размера (4a и видео №2).
Изображение №4
Видео №2: деление капель.
Растяжение и деление капель происходило как следствие сил, вызванных тепловым градиентом на границе раздела газ-вода. Дополнительные опыты с применением капель CM-Dex:pLys показали аналогичный результат. Это значит, что деление капель происходит вследствие воздействия именно физических сил, а не химического состава.
В дополнение к конвекции и капиллярным силам на границе раздела, наличие пузырька газа создает круговорот воды, который также может влиять на поведение и свойства коацервата. Например, циклы «мокрый-сухой» могут привести к накоплению, сушке и регидратации молекул на поверхности. Предыдущие исследования показали, что нагретый газовый пузырь, контактирующий с холодной поверхностью внутри тепловой ловушки, будет имитировать микрожидкостный круговорот воды. Чистая вода из основного раствора будет испаряться на теплой стороне и конденсироваться на холодной. Эти капли воды увеличиваются в размерах и снова попадают в раствор. Испарение, конденсация воды и повторное попадание в объемный раствор приводит к уменьшению (испарение) и увеличению (осадки) высоты границы раздела.
Дабы проверить этот эффект в рамках исследования, ученые загрузили капли CM-Dex:PDDA в тепловую ловушку с температурным градиентом 34 °C (теплая сторона) / 15 °C (холодная сторона).
Микроскопия (4b и видео №3) показала, что капли коацервата накапливались на границе раздела газ-вода и прилипали к теплой поверхности ловушки по мере того, как высота границы раздела уменьшалась из-за испарения воды. Это приводило к тому, что накопленные коацерваты переходили в квазисухое состояние на поверхности.
Видео №3: фрагментация капель.
Сухие полимеры (стрелки на 4b) позже были регидратированы, а пертурбативные потоки, вызванные осаждением воды, приводили к их фрагментации. Образовавшиеся дочерние капли меньшего размера падали в объем и циркулировали в конвекционном потоке. Следовательно, эффект циклов «мокрый-сухой» все же вносят свой вклад в динамику капель на границе раздела газ-вода. Как и ранее, дополнительные опыты с различными вариантами коацервата показали аналогичный результат, подтверждая, что химический состав роли не играет, в отличие от физических процессов.
Недавние исследования показали, что компартментализация путем коацервации или гидрофобный эффект жирных кислот может вносить свой вклад в накопление и регулирование активности РНК. Ученые решили проверить, влияет ли динамика, наблюдаемая в тепловой ловушке, на дисперсию CM-Dex, pLys и РНК.
Для этого были приготовлены дисперсные системы CM-Dex и pLys с РНК и без РНК (2e). Для изучения совместной локализации между РНК, CM-Dex и pLys использовалась двухканальная флуоресцентная визуализация. РНК была помечена ROX (карбокси-X-родамин), тогда как 0.1% компонентов коацервата (CM-Dex или pLys) содержали метку FITC (5а).

Изображение №5
После загрузки дисперсной системы CM-Dex и pLys с РНК в камеру для образцов визуализация показала, что предварительно сформировались маленькие капли коацервата (размером < 15 мкм) в основной колокализованной РНК. Микроскопия показала, что еще до применения температурного градиента капли были богаты на РНК и pLys (CM-Dex также был, но в меньшей степени выражен). Это указывает на то, что РНК сильно конкурирует с CM-Dex за образование капель с pLys.
При наличии температурного градиента (34 °C / 15 °C) наблюдались те же явления, что и описанные ранее: капли коацервата накапливались на границе раздела и сливались воедино.
Капли на границе раздела были больше и содержали все три компонента коацервата (CM: Dex, РНК и pLys; 5a и видео №4), а капли в основном объеме внутри полости оставались маленькими и богатыми на РНК и pLys (5d-5f).
Видео №4: CM-Dex и RNA на границе раздела газ-вода.
Это наблюдение, скорее всего, связано со способностью тепловой ловушки управлять накоплением РНК, pLys и CM-Dex в растворе на границе раздела газ-вода и вызывать обогащение капель этими тремя компонентами.
Количественная оценка размеров капель на границе раздела после применения температурного градиента в течение 1 часа показала, что в присутствии РНК средний размер коацервата упал с 69 ± 31 мкм до 25 ± 9 мкм (5g-5j).
Результаты показывают, как тепловая ловушка может удерживать капли коацервата в неравновесном состоянии, обеспечивая энергетически невыгодные взаимодействия на границе раздела. Это позволяет формировать и выбирать две разные популяции капель внутри полости с разными физическими свойствами и различным составом. Результаты также показывают, что химический состав капель коацервата будет влиять на их фенотип с меньшим размером капель для увеличения концентрации РНК.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Еще в 2018 году авторы рассмотренного нами сегодня труда выяснили, что простая РНК демонстрирует активность в безмембранных микрокаплях, создавая подходящую химическую среду для образования жизни. Те эксперименты проводились в водной среде, где конкурирующие силы были уравновешены. Но для клеток нужна среда, где они могут непрерывно делиться и развиваться.
В нынешней же работе ученые показали, что такой средой может быть газовый пузырь внутри полости. Созданные в лаборатории условия тепловые ловушки напоминают условия, которые можно было наблюдать на ранних этапах развития планеты, когда пористые породы в воде в непосредственной близости от вулканической активности были частично нагреты. Созданная тепловая ловушка представляла собой водосодержащую полость с газовым пузырем и температурным градиентом, когда имеется разница в температуре противоположных стенок камеры. Туда же помещались микрокапли дисперсной системы с определенным составом.
Опыты показали, что физические процессы, протекающие внутри ловушки, способствовали перемещению капель ближе к границе раздела газ-вода, где они подвергаются делению и/или фрагментации. Следовательно, подобные условия могут вполне быть одним из вариантов формирования безмембранных протоклеток миллиарды лет тому назад. Помимо деления и эволюции было обнаружено, что в результате теплового градиента сформировалось несколько типов протоклеток с различным химическим составом, размером и физическими свойствами. Другими словами, температурного градиента было достаточно для запуска процесса эволюционного отбора.
Понимание того, какие именно условия были необходимы для зарождения жизни, позволяют лучше понять весь ее эволюционный путь протяженностью в миллиарды лет. Конечно, остается немало вопросов, но они всегда были и всегда будут. Но каким бы сложным ни был путь поиска ответов, жажда знаний и здравое любопытство человека всегда приведут к финишу.
Благодарю за внимание, отстаивайтесь любопытствующими и отличных всем выходных, ребята! :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Комментарии (6)

zxcvbv
11.12.2021 08:54Официальный старожил-рекордсмен (чей возраст был подтвержден документально) Жанна Кальман прожила 122 года (1875-1997).
Возраст данной мадам последние годы всё больше подвергается сомнению. Тест ДНК пока что так и не проведен.

Overphase
11.12.2021 21:54Да, тоже хотел заметить про "Жанну" Кальман. Сомнения звучат весьма обоснованно, подробнее было именно на Хабре https://habr.com/ru/post/432462/
vgogolin
11.12.2021 12:02+1Газ-вода... Компартментализация... Клетки... Люди. В художественном смысле, получается, мы все - капельки дождя скучавшего на камне под солнцем?

zhabr
если человеческий организм меняется полностью за 7-10 лет, то 122-летняя мадам поменялась раз 10 точно
Radisto
Зубы, хрусталик, еще что-то кажется не меняются. А с возрастом много чего меняется не полностью