
Память является ключевым элементом в функционировании нашего мозга, обладая множеством разновидностей, каждая из которых играет свою особую роль. Существуют декларативная, эмоциональная, сенсорная, временная и долговременная память, и это лишь начало списка. Но давайте подойдем к этому вопросу с инженерной точки зрения: какие типы памяти можно выделить, если анализировать саму природу биологической памяти и механизмы, на которых она строится? Какие методы позволят нам воссоздать эти уникальные процессы? - Давайте разберемся вместе. Конечно биологическая память неразрывно связана с механизмом обучения - подкрепления, которые мы тоже разберем, а также выясним чего же не хватает правилу Хебба. И сделаем мы это максимально наглядно и доступно.
С точки зрения природы биологической памяти на клеточном или молекулярном уровне, её можно разделить на три вида: моментальная, кратковременная и долговременная. Да, всего три механизма памяти в биологическом мозге которые обеспечивают многообразие видов памяти в зависимости от её применения в различных задачах и обстоятельствах.
Моментальная память (оперативная)
Моментальная память - эта та память которая формируется и длиться практически мгновения и хранит информацию которая находится в фокусе внимания. Например, пока человек произносит слово или предложение, он должен помнить, а какое собственно слово он произносит в данный момент, какой контекст предложения, или какое действие непосредственно сейчас совершается. Такая память должна формироваться моментально, а значит не может основываться на каких-либо метаболических изменения в нейронах или в синапсах. Просто на это не хватит времени. Такая память это результат работы механизма реверберации и латерального ингибирования.
Реверберация это циклическая передача возбуждения между различными областями мозга. Рефлекторная дуга не представляет собой последовательный прямой путь передачи возбуждения от заголовка рефлекса (рецепторов) к моторным единицам, как можно представить. На самом деле на всём протяжении рефлекторной дуги возникают кольцевые передачи возбуждения, обозначая на некоторое время весь путь пройденный возбуждением. Например, возбуждение возникшее светочувствительных клетках сетчатки по зрительному нерву попадает в часть таламуса под названием латеральное коленчатое тело откуда в первичную зрительную кору, между первичной зрительной корой и таламусом существуют обратные связи, благодаря которым возникает устойчивый контур передачи возбуждения между таламусом и зрительной корой. Подобным образом формируются циклические передачи дальше по пути рефлекторной дуги: между первичной зрительная корой (V1) и вторичной зрительной корой (V2), между V2 и V3, между V3 и MT, между V3-MT и ассоциативной корой, между ассоциативной корой и префронтальной, между префронтальной и моторной. Это упрощенно, но схематично это так происходит. Эти реверберации скоротечны, цикличность сходит на нет из-за ограниченного ресурса клеток участвующих в реверберации, в синаптических окончаниях ограниченное количество нейротрансмиттера подготовленного для высвобождения в синаптическую щель.
Но всё же такие контуры передачи возбуждения можно рассматривать как механизм моментальной памяти.
Чтобы реверберации использовать в роли память нужен еще механизм латерального ингибирования, или по простому боковое торможение. Латеральное ингибирование это типичное для нервных систем схема организации сети, когда нейрон или структурная единица коры колонка оказывает на соседние в окружении нейроны или колонки тормозящее, ингибирующее воздействие. В связи с чем возникает некая конкуренция между соседними нейронами или колонками. Такие конкурирующие на одном поле колонки можно выделить в группы - гиперколонки или модули. В биологии обычно у модулей нет четких границ, но инженерный подход нам позволяет выделять подобные структуры.
С помощью редактора cogFlux мы можем создать схемы ячейки памяти основанной на реверберации.
Версия на спайковой версии:

Такой элемент памяти можно назвать реверберационный модуль, в нём может быть активен только один реверберационный контур. Мы можем оценить размеры таких модулей в мозге, только по работе Дэвида Хьюбела и Торсена Визела в которой они инвазивно исследовали первичную зрительную кору кошки (“Глаз, мозг, зрение.” 1990 - Хьюбел Д.). В своей работе они указывают что модуль или гиперколонка содержит около тысячи колонок коры (учитывая оба глаза), то есть только одна из пятисот колонок с общим рецептивным полем будет максимально реагировать на предъявленный раздражитель. Можно сказать, что реверберационные модули первичной зрительной коры кошки имеют пятьсот контуров.
Версия на тонических нейронах (5 реверберационных контруров):

Тонические нейроны - это условная математическая модель нейрона, активность которого обозначает частоту генерируемую условным нейроном импульсов и обозначается вещественным числом от 0 до 1.
С элементом затухания (имитация механизмов привыкания и усталости биологических нейронов):

Если модуль или реверберационная ячейка памяти будет иметь семь контуров, то можно сказать что она хранит три бита информации, то есть может иметь восемь состояний.
Преимущества такой памяти - это скорость формирования и чтения памяти, не требуется никаких структурных изменений в сети нейронов. Недостаток - невозможность длительно хранить информацию таким методом, вновь поступившая информация извне полностью стирает предыдущую.
Но на помощь может прийти механизмы кратковременной памяти.
Кратковременная память
Кратковременная память - это память которая длиться ограниченное время от нескольких минут, часов, дней и иногда недель. Информация хранящиеся в кратковременной памяти может исчезнуть по истечении времени если к ней не обратиться.
Основана такая память на механизме NMDA-рецепторов и долговременной потенциации. Долговременная потенциация - это сохранение некоторое время нейроном или нервной тканью высокой возбудимостью или чувствительностью к возбуждению, после некоторого интенсивного воздействия возбуждением. Впервые долговременную потенциацию обнаружили в гиппокампе случайно воздействовав на его ткань токами высокой частоты.
Внимание! Возможна путаница: память кратковременная, а потенциация долговременная (Long-term potentiation). Исследователи обнаруживающие эффект назвали его так, потому что изменения чувствительности обычно происходило на короткие промежутки времени, доли секунд (PTP), но если воздействовать на нейрон множеством импульсов в короткий промежуток времени, то чувствительность менялось надолго относительно этого времени, на часы или дни, то есть на длительно. Но часы и дни в отношении долговременной памяти которая может длиться фактически всю жизнь - это кратковременный интервал, поэтому эффект долговременной потенциации связан с кратковременной памятью.
Эффект долговременной потенциации возможен благодаря особым типам рецепторов в синапсах нейронов - NMDA-рецепторов. “Ключик” к NMDA-рецептору это глутамат, но обычно ионный канал этого рецептора закрыт ионом магния (Mg+) и фактически даже при связывании с глутаматом рецептор не работает. Для того, чтобы он заработал нужно “выбить” так называемую “магниевую пробку” с помощью большой концентрации ионов кальция (Ca2+) под мембраной нейрона. Как мы знаем концентрация кальция под мембраной нарастает при прохождении потенциала действия по мембране, но зачастую необходимо прохождения нескольких импульсов возбуждения по клетке, чтобы сформировалось достаточная концентрация ионов кальция. Получается синапс имеет запас заблокированных рецепторов, но после нескольких активаций, многие заблокированные рецепторы включаются в работу и соответственно чувствительность нейрона возрастает. Со временем ионы магния возвращаются в NMDA-рецепторы и чувствительность нейрона, если не произошло других метаболических изменений, возвращается в исходное положение.

NMDA-рецепторы в мозге распределены неравномерно. Наибольшую концентрацию эти рецепторы имею в области где был обнаружен эффект долговременной потенциации - гиппокампе, также NMDA-рецепторы в большом количестве имеются в коре и базальных ганглиях. В роли памяти здесь выступает изменение в чувствительности нейрона, не отдельных синапсов, а всех синапсов в которых присутствуют NMDA-рецепторы. Так как потенциал действия проходит по всей мембране нейрона, концентрация ионов кальция возрастает для всей мембраны, соответственно изменения чувствительности характерно для всего нейрона.

Повышению чувствительности нейрона даёт преимущество в конкуренции между колонками в модуле или реверберационной ячейке памяти. По сути кратковременная память это расширение моментальной памяти. Чувствительность нейронов колонки участвующей в достаточно продолжительной реверберации может резко увеличится благодаря эффекту долговременной потенциации. Тогда в следующий раз появится преимущество в конкуренции у данного контура реверберации.
Таким образом моментальная память может перетечь в кратковременную память. Помимо самой функции памяти, кратковременная память еще и выполняет некоторые вычисления по распознаванию. Информация которая будет представлена не в полной или зашумленной форме будет распознана и восстановлена в пользу недавно представленной. Реверберационные контуры которые были задействованы в течении дня, будут преимущественно активироваться в продолжении этого дня и дальше.
Открытие долговременной потенциации породило большое количество исследований направленных на изучение пластичности синаптических связей, в частности важным моментом была проверка и уточнение правила Хебба. В теории Хебба не было указаний на то, каким образом влияет временной фактор на синаптическую пластичность. Уже 1997г. был зафиксирован эффект который был противоположен долговременной потенциации (LTP) - это долговременная депрессия (LTD), он связан с снижением возбуждающего посинаптического потенциала (EPSC) при многократной активации постсинаптического нейрона перед пресинаптическим.

Впоследствии была выявлена закономерность которую назвали асинхронный Хебб или STDP (spike-timing-dependent plasticity .англ., пластичность, зависящая от времени импульса). Причем в исследованиях обычно использовались тоже место в гиппокампе, что и при открытии LTP.
В дальнейшем многие исследователи пытались повторить столь элегантный результат с STDP, но вместе с этим получало множество вариантов временных зависимостей, результат зависел от множества условий проводимого эксперимента, от выбранной ткани и её зрелости, от частоты и наличия ингибирующего фактора. Механизм возникновения эффекта долговременной депрессии (LTD) до конца не известен, он не может быть объяснён наличием NMDA-рецепторов. Но, тот факт, что LTD возникает в случае исследований in vivo, где нет изоляции нервных схем от ингибирующего фактора (пресловутое латеральное ингибирование), то возможно LTD имеет скорее внешнюю природу, усиление тормозящего фактора соседних нейронов.

Но все эти эксперименты объединяет то, что пластичность изучалась на основе эффекта LTP, что на мой взгляд не совсем корректно. Во-первых, долговременная потенциация основанная на NMDA-рецепторах, которые активируются при многократном прохождении потенциала действия, потенциал действия распространяется по всей мембране, включая самые удаленные дендриты и окончания, соответственно пластичность изменяется ни на отдельном синапсе, а для всей мембраны в целом, это скорее свойство нейрона, а не отдельных взаимосвязей. Во-вторых, на оси ординат этих зависимостей отмечается возбуждающий постсинаптический потенциал (EPSC) - эта характеристика отражает изменения только на мембране постсинапса, по ней мы ничего не можем сказать о качественных изменениях пресинапса. В-третьих, NMDA-рецепторы это не повсеместное явление, их много, но пластические изменения происходят и в областях и нейронах где их совсем нет.
Долговременная потенциация является важной составляющей механикой кратковременной памяти, но этот эффект не приводит к перестройке синаптических связей (требуется лишь удалить один ион из ионного канала), с этим уже связаны механизмы долговременной памяти.
Долговременная память
Долговременная память это память которая может сохранятся на неограниченный срок. Фактически такая память требует структурных изменений в нейронной сети. Такими изменениями являются перестройки нейронных связей, изменения в размерах синаптических шипиков - утолщений на кончиках синапсов. Дело в том, что размер синаптического контакта определяет количество везикул с нейромедиатором в синапсе подготовленным к выбросу в синаптическую щель. Чем больше размер синаптического шипика тем больше разовая порция выброса нейромедиатора. Также размер синаптического контакта определяет количество рецепторов размещенных на постсинаптической мембране, что определяет общий вклад синапса в поляризацию мембраны нейрона при активации синапса. Размер синапса характеризует его силу или вес. Остается разобраться по каким правилам и как происходит структурная перестройка синапсов.
Правило Хебба
Уже упомянутое правило Хебба можно свести к очень простому тезису: “Если нейроны срабатывают близко по времени, то стоит усилить между ними связь.” И это правило должно проявляться именно в изменении связей между парой нейронов, а не в изменении свойств отдельного нейрона как это проявляется в случае c LTP. В этом плане мы должны фиксировать качественные изменения в пресинаптических шипиках, такие изменения как размера или количество везикул в нём. И это должны быть наблюдения длительного периода с сопоставлением времени срабатывания пре- и пост- синапсов, что выполнить крайне сложно. Главным доказательством верности теории Хебба пока является наличие ассоциативного обучения.
Временная зависимость в правиле Хебба является открытым вопросом, и теоретически оно может иметь различные формы, возможны классическая (симметричный Хебб) и обратная версия этого правила (анти-Хебб), и версии где имеет значение последовательность срабатывания пар нейронов (асимметричное правило). В случае ингибирующего нейрона, все медиаторы в синапсах которого оказывают тормозящее воздействие, правило Хебба по логике теории нейронных ансамблей должно работать в обратную сторону, или по анити-Хеббу.

Симметричность или асимметричность правила Хебба определяется скорее задачами применения сети и нейроном в котором оно обнаруживается. Если в обучении важную роль играет фактор времени или последовательности как задачах управления движением, эффективным будет использование нейронами асиметричного правила Хебба. В задаче получения ассоциативных взаимосвязей лучшим вариантом будет симметричный Хебб.
В обычном, классическом виде правило Хебба не показала свою эффективность в сравнении с результатами распространенного в машинном обучении алгоритма обратного распространения ошибки. И дело здесь не в самом правиле, а том, как его применять и в способе организации сети в котором его следует применять, так как это правило успешно работает в биологических сетях.
Во-первых, правило Хебба лучше работает в сочетании с разреженными данными, то есть данными которые представлены в виде большинства неактивных элементов и единичным количеством активных, такое соотношение позволяет более прицельно настраивать связи между нейронами. Во-вторых, правило Хебба можно развить и внести еще небольшие дополнения которое сильно усиливает его эффективность, это добавление к классическому Хеббу (короткий Хебб) правило длинного Хебба. Классическое правило Хебба работает с единичными спайками, то есть оно применяется по результатам прохождения одного импульса потенциала действия по синапсу. Для применения длинного Хебба требуется набрать некую статистику работы синапса и если синапс используется так, что регулярно возникшая активность у пресинаптического нейрона предшествует, или происходит близко по времени активности постсинаптического нейрона, то следует укрепить связь. И такое укрепление связи будет происходить при каждом прохождении импульса по синапсу.
Такое сочетание короткого и длинного Хебба даёт важные улучшения. Наличие только короткого Хебба приводит нестабильному обучению и постоянной перестройке связей если данные будут нестабильны и зашумленные, а дополнение в виде долгого Хебба сформирует устойчивые связи. С другой стороны без короткого Хебба невозможно переобучение, а также короткий Хебб важен на старте обучения. Подробнее о данной модели можно прочитать в моей прошлой статье (https://habr.com/ru/articles/709350/).
Но, не всё так просто и требуется представление того как всё таки нейрон понимает какой синапс следует модифицировать и как Хебб связан с подкреплением.
Прионная теория долговременной памяти
Нейрон - это биологическая клетка, и как клетка он имеет все важные атрибуты клетки. Как и у большинства клеток организма у нейрона есть ядро, которое располагается примерно в центре клетки. Клеточное ядро - это хранилище “чертежей” всех белков организма, с помощью него можно инициализировать производство необходимых белков в нужном количестве. Рядом с ядром обычно расположена центриоль - органелла от которой начинают свой рост микротрубочки которые составляют каркас для клетки или цитоскелет.
")
Цитоскелет находится в постоянной динамике. Микротрубочки постоянно разрушаются и формируются. Они растут от отрицательного центра - центриоли, к положительным кончикам у мембраны.
Цитоскелет - это не просто каркас, но и важная транспортная система. По ним двигаются специальные моторные белки - кинезины. Кинезины - это буквально ходячие белки.

Один конец кинезина сцеплен с микротрубочкой и шагает по ней, другой связан с грузом, обычно это пузырёк мембраны с необходимыми молекулярными механизмами на ней (рецепторами, ионными насосами) или внутри пузырька может переноситься необходимые молекулы веществ (гормоны, ферменты), например, в случае нейрона - молекулы нейромедиатора.
Моторных белков существует несколько видов. Есть те которые двигаются от центра клетки к положительному концу микротрубочки и приносят мембране необходимый груз (кинезин), есть те которые двигаются от мембраны в центру унося обратно мусор и лишние белки (динеин).

В нейронах можно выделить разные кинезины которые относят грузы по дендритам и соме и кинезины которые переносят грузы по аксону и его терминалям. Мы помним, что длина некоторых аксонов в организме достигает 1 метра, а грузы для пре- и пост- синапсов могут сильно отличаться, чтобы не таскать в пустую ненужный груз на такое расстояние эволюция разделила эти виды кинезинов. Поэтому в нейронах в которых нету явного внешнего разделения на дендриды и аксон, к примеру, маленькие звездчатые нейроны аксон которых не имеет миелиновой оболочки, можно определить отросток аксон по типу кинезинов премещающихся в нем. Это можно сделать с помощью специальных флуоресцентных технологий. Скорость кинезина в аксоне достигает 500 мм в сутки.
Как мы видим в этой транспортной системе отсутствует адресность. Сформированные белки у ядра клетки будут разносится ко всем её частям, да, есть разграничение между аксоном и дендритами, но у аксона может быть сотни терминалей и тысячи синаптических окончаний. Как система клетки определяет какому синапсу нужно применить те белки которые были транслированы в её центре? - На помощь могут придти не менее интересные белки прионы.
Белки прионы были обнаружены когда искали возбудитель болезни куру. Было невероятным, то что существует белок способный к самокопированию без участия нуклеиновых кислот. Известно, что нормальная форма белка имеет какую-то важную функцию в работе нервной системе, но до конца функции прионов в организме не известны. Также существует патогенная форма белка, которая и является возбудителем болезни куру, этот белок просто скручен по другому и при контакте с другим нормальным белком патогенный может изменить его конформацию с нормальной на патогенную. Помимо прионов в организме существуют белки которые контролируют и регулируют их размножение, но эти белки не распознают ненормальную форму приона. Что приводит к тому, что патогенная форма приона может неконтролируемо размножаться. Многие связывают патогенную форму прионов с различными нейродегенеративными заболеваниями. Появление такого патогена в организме может произойти не только в случае ошибки трансляции белка или генетической ошибки, но и через пищу, как в случае с болезнью куру. Прионная теория памяти поясняет зачем нейронам нужен столь опасный белок.
Правильная форма белка приона обладает важными качествами чтобы быть молекулой памяти. Это возможность запустить контролируемый процесс самокопирования, фактически с небольшого количества молекул добиться высокой локальной концентрации прионов и поддержания этой концентрации достаточно продолжительное время.

Представим систему из трех нейронов, где мы рассмотрим два пресинапса. Одни синапс соединяет пару нейронов которые сработали совместно в короткий промежуток, второй синапс соединяет нейроны которые совместно не срабатывали. Соответственно в первом синапсе согласно правилу Хебба следует произвести структурные изменения по увеличению его силы, по сути увеличить его размер. Но подобные изменения требуют значительного времени, и чтобы не потерять информацию о требуемом изменении в этом случае возможно запустить работу приона, который локально только в пределах данного синапса увеличит свою концентрацию на некоторое время, тем самым служа некой меткой. Таким образом возможно пометить синапсы которые требуют изменений до их окончания. Требуемые строительные материалы будут созданы у центра клетки, доставлены с помощью транспортной системы во все синаптические окончания, но будут применены только в тех синапсах где есть прионная метка.
Подкрепление и пластичность
Известная истина: мозг пластичен. Пластичность мозга - это способность его перестраиваться при необходимости. Пластичность эта неравномерна, есть области мозга в которых высокая степень пластичности сохраняется даже в очень зрелом возрасте. К примеру, нейрогенез, то есть создание новых нейронов, возможен только в некоторой части гиппокампа и в области ответственной за обоняние откуда новорожденные нейроны могут мигрировать и в другие соседние области. К механизму пластичности относится не только нейрогенез, но механизмы перестройки синаптических связей. Хотя термин пластичность больше применим к мозгу и нервной системе, мы можем пластичностью охарактеризовать отдельный нейрон как составную часть системы. Пластичность для нейрона это мера скорости с которой происходят изменения в его синаптических связях. К примеру, если ядро нейрона траслирует очень малое количество белков для строительного материала синапса, то даже при наличии прионной метки до конца действия её срока изменения в синапсе произойдут в малой степени или вообще не произойдут. В случае же когда ядро нейрона транслирует очень большое количество строительного материала, изменения в синапсах происходят быстро и с большой степенью.
Одним из важных элементов подкрепления является медиатор дофамин и дофаминергическая система мозга. Дофаминовые нейроны, медиатором которых является дофамин расположены в небольших областях мозга вентральная область покрышки (ventral tegmental area) и черное вещество (substantia nigra). Аксоны этих нейронов распространяются и тянутся на большие области коры, гиппокампа, таламуса и гипофиза. Эти аксоны имеют множество разветвлений - терминалей, и варикозных утолщений которые при активации нейрона выбрасывают в межклеточное пространство медиатор дофамин. Дофаминовые рецепторы нейронов метаботропные, то есть они напрямую не влияют на поляризацию мембраны, а встроены в мембрану таким образом, что один конец рецептора который может сцепляться с молекулой дофамина находится снаружи клетки, другой конец находится под мембраной внутри клетки и обычно сцеплен со специальной молекулой лигандом, которая при активации рецептора отсоединяется. Свободный лиганд может активировать ионный канал или запускает цепочку реакций которая изменяет метаболизм клетки. То есть медиатор дофамин оказывает модулирующее действие на целевой нейрон. Модулирующее действие может быть различного свойства, к примеру, мы можем заставить ядро клетки на короткий срок создать больше строительного материала для того, чтобы произвести изменения в синапсах.

Представим схему из трех нейронов. Одни синапс соединяет пару нейронов которые сработали совместно, соответственно срабатывает правило Хебба и данный синапс помечается прионной меткой. В обычном состоянии этот нейрон имеет очень низкую пластичность и по истечении срока действия метки, если не произойдет никаких изменений, перестройки в синаптической связи произойдут незначительные. Но если до истечения срока действия прионной метки произойдёт подкрепление (reward) и на клетку подействует медиатор дофамин, то это повысит на короткий срок пластичность нейрона и запланированные изменения в синапсах произойдут быстро и в полном объеме.
Этот механизм объясняет как в случае обучения животных происходит ретроспективность подкрепления. Награда может действовать на события и навыки полученные некоторое время назад подкрепляя их.

Представьте нейронную сеть, которая имеет некоторое количество связей и соответственно значение весов для каждой настраиваемой связи - параметры. После акта обучения основные параметры сети не изменяются, а изменения происходят в некой копии параметров сети - временные целевые параметры. Условно эти целевые параметры хранятся 20 минут. Если акт обучения остался без подкрепления, то через 20 минут целевые параметры вернутся к значениям базовых параметров и информация об акте научения будет утеряна за ненужностью. Если в течении этих 20 минут сеть получила подкрепление, то базовые параметры принимают значение целевых параметров.

Такую схему научения и подкрепления не получится применить к классическим конекционным сетям, так как при каждом акте обучения в сети затрагиваются каждый нейрон и каждый параметр, который немного изменяет под действием алгоритма обратного распространения ошибки. Сети нашего мозга работают по нейро-детекторному принципу и разреженности. Во-первых, это дает практически автоматическое разбиение информации поступающей в сеть на некие метки - якоря: объекты, субъекты, действия, события. Во-вторых, различные акты обучения будут затрагивать разные нейроны, что позволяет разделять полученные навыки даже если они давались в близкое время.
Конечно, ключевую роль в научении будет играть временная близость события и подкрепления. Ярким примером здесь служит суеверное поведение (superstitious behavior). Когда испытуемые голуби в ящике Скиннера, имели случайное пищевое подкрепление и заучивали некоторые движения и действия случайно предшествовали подкреплению, но не были никак с ним связаны. К примеру, поднимали одно крыло или клевали в определённом углу камеры. Причем регулярно совершали эти суеверные действия полагая, что так они получают награду.

Эксперименты Скиннера говорят нам, что живое существо всегда будет искать и связывать события которые предшествовали подкреплению, даже если этих связей объективно нет. Если такие события есть, то предпочтение будет у события для связи с подкреплением который будет ближе по времени и с высокой корреляцией.

Представим, что существуют события A, B, C которые предшествуют возникновению подкрепления D. Установлено, что событие A является причиной D хоть и является самым далеким событием отстающим по времени, а событие B коррелирует с D в 50% случаях, событие C коррелирует в 1% случаях. Если при первом акте все события сработали перед подкреплением, то максимально связь усилится между событием C и D. В последующих актах обучения, так как событие C будет редко предшествовать подкреплению, то связь C и D будет деградировать, за счет гетеросинаптической конкуренции. Связь между событиями A и D будет только усиливаться. Также связь будет достаточно крепка и устойчива между событиями с высоким уровнем корреляции как между B и D. Полностью избавиться от суеверного поведения данная система к сожалению не может.
Связь кратковременной и долговременной памяти. Консолидация
Эффект переноса памяти из временной в долговременную называют консолидацией памяти. Существуют два важных условия для консолидации памяти - это повторение информации, иногда требуется неоднократное повторение и эмоциональная значимость информации, если информация имеет высокую значимость для организма, то такая информация может сохраняться сразу в долговременную память.
Но по существу не происходит непосредственного переноса информации из систем временной памяти в системы долговременной, как это может происходить в компьютерных системах, дело в том, что эти механизмы работают независимо и параллельно. Одна и та же информация попадает в обе системы параллельно. Система временной памяти гиппокамп и другие сети нейронов обладающие NMDA-рецепторами, с первых моментов могут реагировать на новую информацию и фиксировать её, но как известно NMDA-рецепторы со временем восстановят свои исходные состояния, утратив информацию. Системы же долговременной памяти отреагирую на новую информацию только прионными метками, которые могут исчезнуть через некоторое время, если информация не окажется эмоционально подкрепленной дофамином, норадреналином или окситоцином. И небольшая пластичность позволяет даже не подкрепленной информации при многократном повторении сохраниться.
В пользу этого, может служить пример с пациентом H.M. (Генри Молисоном) которому Уильям Сковилл упешно удалил (разрушил) обе половины гиппокампа. Впоследствии Генри утратил способность к временной памяти, при утрате внимания с какого-либо действии он переставал помнить что происходило до этого, его воспоминания о событиях были зафиксированы только до 1953г. При этом он мог формировать новые моторные навыки при многократном повторении конкретных действий. Подробней о механизмах консолидации можно прочитать в моей прошлой статье (https://habr.com/ru/articles/397377/).

Многое указывает, что сон может способствовать переходу информации из кратковременной памяти в долговременную. Можно предположить причины этого, но в вопросе снов и сновидений существует множество неисследованных моментов. В момент сна мозг переходит некоторый иной режим, и префронтальная кора в некоторой степени уменьшает свой контроль над подкорковыми эмоциональными центрами. В этом случае активизируется некоторый контур реверберации называемый круг Пейпеца в котором вовлекаются эмоциональные центры и гиппокамп и некоторые области коры. При такой реверберации с большей вероятностью будут активированы контуры помеченные долговременной потенциацией, то есть во сне события дня особенно которые имели эмоциональный окрас будут проигрываться в кратковременной памяти. Но фактически повторение информации во сне эквивалентно повторению информации при бодрствовании для системы долговременной памяти. И в процессе сна и работы ревербераций круга Пейпеца, такое повторение может происходит многократно, но к сожалению этот процесс неконтролируем и мы не можем выбрать перед сном информацию для такого прогона. С другой стороны сон это время очистки мозга, в мозге нет лимфатических узлов и лимфы, и альтернатива этому - это сужение сосудов мозга во время сна, которое приводят и к оттоку жидкостей содержащих ненужные продукты жизнедеятельности и прочий сор, а также это время избавления от накопившихся незадействованных прионных меток. Сложный механизм сна приводит в порядок информацию полученную за день, убирая малозначимое и укрепляя эмоционально значимое.
Специализированная память
Итак, у нас есть три механики памяти: реверберации, NMDA-рецепторы и прионные метки с последующей пластичной перестройкой синапса. Эти механики позволяют оперировать памятью непрерывно переходя от одной формы к другой. Фактически нет четких границ того какая из систем в данный момент является хранилищем информации, это всё работает как единая система. Если говорить о других формах классификации памяти, то некоторые мы можем отнести к конкретным механикам. Если говорить о процедурной памяти, та которая хранит неосознанную информацию, в частности “мышечную память”, какие-либо моторные навыки, то она фактически может миновать моментальную и кратковременную память, а сразу задействовать механику прионных меток, и если навык будет подкреплён - это поможет быстро усвоить навык, либо потребует множество повторений. Соответственно моторные навыки будут локализоваться в моторной и премоторной коре, также в базальных ганглиях и мозжечке. Та информация которая воспринимается осознанно она обязательно проходит через моментальную (оперативную) и кратковременную память. Осознаваемая информация храниться в так называемой декларативной памяти. Подвид декларативной памяти, есть семантическая память в которой хранятся различные факты о мире, например, “трава-зеленая”, “Солнце - это звезда” и это не только информация которая выражается речью, еще пример - расположение элементов лица человека, форма тела и так далее. Прежде чем сохраниться в долговременной памяти семантическая информация должна быть осознана и зафиксирована в кратковременной памяти, и если она имеет ценность будет уже сохранена в долговременной памяти. Локально такая память фактически распределена по всей коре больших полушарий.
Эпизодическая память также является декларативной, то есть осознаваемой, но её формирование прежде всего сконцентрировано в гиппокампе и соседних областях коры. Гиппокамп имеет специализированные области отвечающие за ментальную карту, фактически информацию о том, где находится в данный момент организм, а также где находится точки интереса на этой карте, границы и другие субъекты. Рядом с ментальной картой обнаруживаются и клетки времени, которые дают представление о текущем относительном времени. Фактически гиппокамп и система кратковременной памяти позволяют отвечать на вопросы “Что? Где? Когда?”. Если описанный кратковременной памятью эпизод окажется ценным и важным, то он сохраняется в долговременной памяти в виде автобиографической памяти в семантической форме, но уже в коре.
В прионной модели памяти возможен только одна форма сигнала - положительное подкрепление, отрицательной формы нет, не существует команды “удалить все прионные метки” . Всё многообразие эмоций используют одну эту форму, в случае получения удовольствия, получения награды - получаем положительное подкрепление. И страх, боль в случае их избегания или окончания тоже приводит к положительному подкреплению, информация запоминается, это нужно, чтобы при успешном уходе от опасности, или избежания боли в следующий раз организм был бы более успешен в этом. Вместе с информацией сохраняется и репрезентативное представление текущего эмоционального состояния, что придаёт памяти различный эмоциональный окрас, отсюда термин эмоциональная память.
Еще одна специализированная память и область мозга - это мозжечок. Мозжечок очень древняя область мозга, которая зарождается очень рано в результате совершенствования произвольных движений и усложнения структуры управления телом. Мозжечок хранит память о точных коротких интервалах времени для управление движением. В мозжечке также в некоторой доли присутствую клетки с NMDA-рецепторами, а значит некоторую информацию он может фиксировать достаточно быстро в кратковременной памяти.
Вместо заключения
Становится ясно, что достичь сильного ИИ возможно различными методами и технологиями. Большие Языковые Модели (LLM) и развитие подобных технологий позволят создать интеллект или даже разум некоторого нового качества совершенно отличного от человеческого. В котором не будет места знаниям о биологии человеческой памяти или сознания. Но я верю и увлечен идеей создания интеллекта основанного на биологических принципах организации памяти, сознания и эмоций. Поэтому изучая нейрофизиологию я сжимаю, упрощаю и систематизирую информацию, чтобы она стала еще более доступна для понимания и реверс-инжиниринга. Хоть этот альтернативный путь возникновения искусственного интеллекта и не имеет столь мощного результата и интереса в сравнении с LLM, но возможно в будущем это будет спасительная веточка для существования Человечества.
О своей работе я делюсь на страницах habr, а также существует ненавязчивый телеграмм-канал (https://t.me/neuro_cyber) и группа в ВК (https://vk.com/neuro_cyber).
Комментарии (2)

phenik
18.04.2024 20:04+1Моментальная память (оперативная)
В классификациях такой нет, что имеется в виду? Близко по смыслу иконическая и рабочая память, которая ограничена. Последнюю иногда называют оперативной, и считают разновидностью кратковременной. Вот тут еще упоминается мгновенная память, относят ее к сенсорной, как и иконическую. Конечно все классификации условны, меняются со временем. Необоснованное добавление еще некой моментальной только увеличивает неразбериху.
Замечаний к тексту не мало, хорошо что хоть не реверсировали память к транзисторам, как это часто делают физики и технические специалисты) напишу только к этому
Становится ясно, что достичь сильного ИИ возможно различными методами и технологиями. Большие Языковые Модели (LLM) и развитие подобных технологий позволят создать интеллект или даже разум некоторого нового качества совершенно отличного от человеческого. В котором не будет места знаниям о биологии человеческой памяти или сознания.
Почему отличного от человеческого? ЯМ, как и любые ИНС построены на сетях формальных нейронов, которые моделируют важную суммативную функцию биологических нейронов. В целом ЯМ моделируют, худо-бедно, ассоциативное мышление и память человека, здесь не плохо показано. Из-за такой ограниченной модели все проблемы ЯМ связанные с так называемыми галлюцинациями, на самом деле фантазиями, и другими немалыми недостатками. У человека, как известно, форм мышления намного больше (здесь подробнее), и соответственно видов памяти, и архитектуре ИНС еще есть куда, и каком направлении совершенствоваться. Для чего современного уровня ЯМ достаточно в практическом плане так это для разработки нового поколения интеллектуальны ассистентов, что собственно и наблюдается.
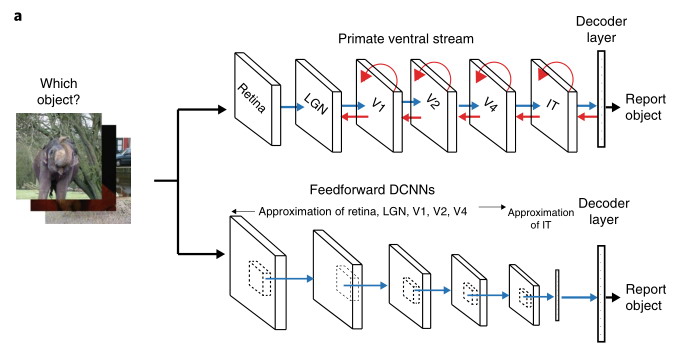
Практически все идеи ИНС почерпнуты в исследованиях биологических прототипов даже если авторы в своих работах не указывают ссылки на соответствующие исследования. Даже такой поборник собственного пути ИИ, как Ян Лекун черпал и черпает свои идеи из работ нейробиологов, сверяет результаты с ними, достаточно посмотреть его записки. Вот схематическое изображение архитектуры глубоких сверточных сетей над которыми он работал в сравнении с вентральным трактом зрительной системы приматов. И даже такая процедура, как обучение с помощью обратного распространения ошибки имеет некоторый ограниченный аналог в биологических сетях, хотя пока и не до конца изученный.
Но я верю и увлечен идеей создания интеллекта основанного на биологических принципах организации памяти, сознания и эмоций. Поэтому изучая нейрофизиологию я сжимаю, упрощаю и систематизирую информацию, чтобы она стала еще более доступна для понимания и реверс-инжиниринга. Хоть этот альтернативный путь возникновения искусственного интеллекта и не имеет столь мощного результата и интереса в сравнении с LLM, но возможно в будущем это будет спасительная веточка для существования Человечества.
Отрадно читать такое, и это не альтернативный путь, а как раз прямой, и он уже давно развивается с использованием нейроморфных технологий. Практически все известные производители компьютерного железа имеют программы их разработки, и даже коммерческие продукты. Эти решения ближе к биологическим прототипам по многим параметрам, включая, по энергоэффективности (беды современных ИНС), импульсному режиму работы, асинхронности, возможным встроенным генераторам паттернов активности, способам обучения, включая непрерывный, и тд. Но сложнее в реализации, и потребуется еще некоторое время, чтобы реализовать все свои возможности. Вероятно вы в курсе эти работ. Это позволит создавать не только интеллектуальных ассистентов, пусть и продвинутых в отдельных областях, но и интеллектуальных агентов, включая воплощенных, которые будут ближе к когнитивной архитектуре (в этом коменте подробнее). Вероятно в перспективе статические решения типа ЯМ займут свою нишу, а в основном будут использоваться нейроморфные решения.
Возможно читали про эту исследовательскую разработку в области импульсных сетей, в кот. для повышения эффективности обучения используется периодическая активация состояния сети являющегося некоторым аналогом сна и консолидации памяти у человека (науч-поп., статья). Несколько курьезно, но демонстрирует возможности технологии, т.к. частично решает проблему катастрафического забывания информации, которая характерна для современных ИНС, хотя над ней усиленно работают.
Удачи в поисках!
{kind=link}
victor_1212
однако, это очень даже интересует, начинаю смотреть весь цикл статей, хотелось бы иметь min информацию об авторе